Контрольная работа: Обмін ліпідів
Контрольная работа: Обмін ліпідів
Обмін ліпідів
План
1. Перетворення ліпідів у харчовому тракті людини.
2. Окислення
гліцерину
3. Окислення
вищих жирних кислот
4. Обмін кетонових
тіл
5. Біосинтез
гліцерину
6. Біосинтез
вищих жирних кислот
7. Біосинтез
тригліцеридів
8. Біосинтез фосфоліпідів
1. Перетворення ліпідів у процесі травлення
Обмін ліпідів– багатоступеневий процес який складається з процесів
травлення в харчовому тракті, всмоктування, транспортування в тканинах
організму, внутріклітинного окислення і біосинтезу.
Поскільки ліпази (ферменти що розщеплюють жири) мають оптимум дії
рН=7.8–8.1, то процеси розщеплення ліпідів сторго локалізовані в кишково–шлунковому
тракті.
|
Відділ харчового тракту
|
Оптимум дії (рН)
|
Процеси
|
| Ротова порожнина |
6.5–7 |
Процеси розщеплення відсутні |
| Шлунок |
1.5–2– |
Розщеплення білків . Звільнення
ліпідів з ліпопротеїдних ком-плексів. |
| 12–пала кишка та відділи тонкого
кишечника |
7.5–8.5 |
Основне місце розщеплення ліпідів. |
|
Зважаючи на гідрофобність ліпідів
та водорозчинність ліпаз, необхідною умовою розщеплення ліпідів є їх
диспергування (подрібнення) та емульгування (збільшення площі поверхні).
Центральною речовиною в процесах розщеплення ліпідів є жовч – в‘язка
рідина світло-жовтого кольору з гірким смаком та специ-фічним запахом. Міститься в жовчному міхурі. До її
складу входять жовчеві кислоти (холева та дезоксихолева), холестерин, вищі
жирні кислоти, три-гліцериди –
фактори емульгування; ферменти і гормони – сприяють розщеп-ленню ліпідів; жовчні пігменти і
продукти розпаду гемоглобіну. Жовч забез-печує перистальтику тонкого кишечника і
нейтралізацію харчового хімусу – фактори диспергування, активацію
ліполітичних ферментів, проникність стінок кишок, транспорт продуктів
ліполізу, бактеріостатичну дію на кишкову мікро-флору, виведення ядів із
організму.
|
Розщеплення тригліцеридів відбувається за допомогою жовчі у 12-палому
ки-шечнику та тонкій кишці під впливом ліпаз, що мають оптимум дії
рН=7.8–8.5, за слідуючою схемою:
CH2–O–C(O)–C17H35 CH2–OH 2C17H35COOH
 CH–O–C(O)–C17H33 ліпаза CH–OH + C17H33COOH CH–O–C(O)–C17H33 ліпаза CH–OH + C17H33COOH
CH2–O–C(O)–C17H35 CH2–OH
    В 12-палій кишці, в
основному, розщеплюються фосфогліцериди під впливом фосфоліпаз (А,В,С,Д).
Останні характеризуються строгою специфічністю своєї дії: Фосфоліпаза
А–каталізує розрив складноефірних зв‘язків між гліцерином та ненасиченими
кислотами, фосфоліпаза В– між гліцерином і насиченою кис-лотою,
фосфоліпаза С–між гліцерином і фосфорною кислотою а фосфоліпаза Д розщеплює тільки
фосфохолін. В 12-палій кишці, в
основному, розщеплюються фосфогліцериди під впливом фосфоліпаз (А,В,С,Д).
Останні характеризуються строгою специфічністю своєї дії: Фосфоліпаза
А–каталізує розрив складноефірних зв‘язків між гліцерином та ненасиченими
кислотами, фосфоліпаза В– між гліцерином і насиченою кис-лотою,
фосфоліпаза С–між гліцерином і фосфорною кислотою а фосфоліпаза Д розщеплює тільки
фосфохолін.
CH2–O–C(O)–C17H35 CH2–O–С(О)–C17H35
CH–O–C(O)–C17H33 фосфоліпаза А, Н2О CH–OН
+ C17H33COОН
       CH2–O–P–O–CH2–CH2–N(CH3)3 СН2–О–Р–О–СН2–СН2–N(CH3)3 CH2–O–P–O–CH2–CH2–N(CH3)3 СН2–О–Р–О–СН2–СН2–N(CH3)3
          HO O HO O HO O HO O
CH2–O–C(O)–C17H35 СН2–ОН
 CH–OH фосфоліпаза В, Н2О СН–ОН + C17H35COOH CH–OH фосфоліпаза В, Н2О СН–ОН + C17H35COOH
      CH2–O–P–O–CH2–CH2–N(CH3)3 СН2–О–Р–О–СН2–СН2–N(CH3)3 CH2–O–P–O–CH2–CH2–N(CH3)3 СН2–О–Р–О–СН2–СН2–N(CH3)3
  HO O HO O HO O HO O
 CH2–OH CH2–OH фосфоліпаза
С, Н2О CH–OH+HO–P–O–CH2–CH2–N(CH3)3 фосфоліпаза
С, Н2О CH–OH+HO–P–O–CH2–CH2–N(CH3)3
 CH2–OH фосфоліпаза
Д, Н2О Н3РО4 + НО–СН2–СН2–N(CH3)3 CH2–OH фосфоліпаза
Д, Н2О Н3РО4 + НО–СН2–СН2–N(CH3)3
Стериди під дією ферменту холестеринестерази розпадаються
до стеринів і вищих жирних кислот згідно схеми:
В результаті ферментативного розщеплення ліпідів, в порожнині тонкої кишки утворюються на 80 % продукти повного
гідролізу (гліцерин, вищі жирні кислоти, азотисті основи і фосфорна кислота) та
20 % продукти часткового гідролізу (ди– і тригліцериди).
Із просвіту тонкого кишечника продукти розщеплення потрапляють у клітини
епітелію . Даний процес проходить із затратами енергії. Необхідно враховувати
що: гліцерин, гліцеринфосфорні кислоти, азотисті основиі фосфорна кислота –
добре розчинні у воді – легко проникають в епітеліальні клітини; вищі жирні
кислоти, холестерин, дигліцериди – погано розчиняються у воді. В епітеліальні
клітини потрапляють у комплексі з жовчевими кислотами, утворюючи водороз-чинні
холеїнові кислоти.
В епітеліальних клітинах тонкого кишечника відбувається
розпад холеїнових кис-лот, синтез специфічних для організму ліпідів та процеси
синтезу транспортних форм для специфічних ліпідів (поскільки більша їх
кількість гідрофобна). Серед транспортних форм специфічних ліпідів виділяють
слідуючі:
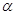 –Ліпопротеїди – рухливий комплекс фосфатидів з білками; –Ліпопротеїди – рухливий комплекс фосфатидів з білками;
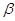 –ліпопротеїди –
рухливий комплекс холестерину та його ефірів з білками; –ліпопротеїди –
рухливий комплекс холестерину та його ефірів з білками;
хіломікрони – рухливий комплекс діаметром 150–200 нм, що
складається з ново-синтезованих (специфічних) тригліцеридів і вітамінів,
зовнішньою обгорткою яких є білкова молекула.
На відміну від вуглеводів котрі відразу потрапляють у кров, транспортні
форми ліпідів розносяться по організму завдяки лімфатичній системі. При
контакті з кровоносними судинами останні під впливом спеціальних ферментів розпада-ються на ліпідний та білковий компоненти. Білковий компонент
током лімфи по-вертається до клітин епітелію тонкого
кишечника де утворює нові транспортні форми, а ліпідна фракція током крові
розноситься до клітин організму де відбу-ваються процеси
їх внутріклітинного окислення.
При виконанні інтенсивної, довготривалої м‘язевої роботи
енергетичні затрати організму покриваються за рахунок енергії окислення
ліпідів.
2. Окислення гліцерину
Проходить у дві стадії за участью різних ферментних систем.
1.Утворення гліцеринфосфорної кислоти за допомогою фермену
гліцеролкінази:
    CH2–OH CH2–OH CH2–OH CH2–OH
  CH–OH АТФ АДФ CH–OH CH–OH АТФ АДФ CH–OH
  CH2–OH CH2–O–P=O CH2–OH CH2–O–P=O
 HO OH HO OH
2. Утворення диоксіацетонфосфату та фосфогліцеринового альдегіду за
допомо-гою НАД-залежних та ізомеразних ферментних систем:
CH2–OH СH2–OH HC=O
CH–OH НАД+ НАДНН C=O ізомераза HC–OH
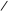               CH2–O–P=O CH2–O–P=O HC–O–P=O CH2–O–P=O CH2–O–P=O HC–O–P=O
HO OH HO OH H HO
OH
гліцеринфосфорна кислота диоксіацетонфосфат фосфогліцериновий альдегід
ОТЖЕ: енергетичний ефект окислення гліцерину до ФГА становить 2
молекули АТФ, а повного окислення до вуглекислого газу і води через ФГА – 22
молекули АТФ. Фосфогліцериновий альдегід – спільний метаболіт для ліпідного і
вугле-водневого обмінів – використовуються для біосинтезу жирів, фосфогліцеридів
і стеридів що проходять у печінці.
3. Окислення вищих жирних кислот
В основі сучасних уявлень про механізм біологічного окислення вищих
жирних кислот лежить теорія запропонована Кноппом у 1904 році. Він
експериментально довів що, окислення ВЖК має характер поступового, циклічного
6-стадійного процесу.
Перші дві стадії – підготовчі, полягають у активації молекул ВЖК з
утворенням їх активованої ацильної форми, ацил–КоА.
Наступні чотири – циклічні, являють собою власне окислення
ацил–КоА з утво-ренням ацетил–КоА, яка доокислюється в циклі реакцій
трикарбонових кислот (циклі Кребса) до вуглекислого газу і води.
Підготовча стадія:
         C13H27 С13Н27 C13H27 C13H27 С13Н27 C13H27
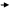 CH2 АТФ Н4Р2О7 СН2 НS–KoA CH2 CH2 АТФ Н4Р2О7 СН2 НS–KoA CH2
  CH2 СН2 CH2 + АМФ CH2 СН2 CH2 + АМФ
 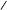   COOH С=О О C=O COOH С=О О C=O
  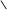 О–Р–О–А S–KoA О–Р–О–А S–KoA
ОН
пальмітинова кислота пальмітил-аденілат пальмітил-КоА
Циклічне окислення:
            C13H27 С13Н27 C13H27 С13H27 C13H27 С13Н27 C13H27 С13H27
CH2 ФАД+ ФАДНН СН HOH CH–OH НАД+НАДНН C=O
 CH2 СН CH2 CH2 CH2 СН CH2 CH2
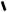    C=O С=О C=O C=O C=O С=О C=O C=O
S–KoA S–KoA S–KoA S–KoA
пальмітил-КоА ненасичена форма оксиформа кетоформа
пальмітил-КоА пальмітил-КоА пальмітил-КоА
  C13H27 CH3 C13H27 CH3
  C=O + C=O C=O + C=O
S–KoA S–KoA
ацильний залишок ацетил–КоА
Ацильний залишок далі доокислюється в 4-х стадійному циклі до повного роз-паду молекули на ацетил–КоА, який доокислюється в циклі Кребса.
Повне окис-лення пальмітинової кислоти з утворенням восьми
молекул ацетил–КоА прохо-дить за сім раз
чотирьохстадійного циклу з виділенням 7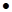 5=35
молекул АТФ. При окисленні однієї молекули ацетил–КоА в циклі Кребса
виділяється 12 моле-кул АТФ, а поскільки з молекули
пальмітинової кислоти утворюється 8 молекул ацетил–КоА, то енергетичний ефект
їх окислення становить 8 5=35
молекул АТФ. При окисленні однієї молекули ацетил–КоА в циклі Кребса
виділяється 12 моле-кул АТФ, а поскільки з молекули
пальмітинової кислоти утворюється 8 молекул ацетил–КоА, то енергетичний ефект
їх окислення становить 8 12=96
молекул АТФ. Загальний енергетичний ефект окислення однієї молекули
пальмітинової кислоти становить 96 12=96
молекул АТФ. Загальний енергетичний ефект окислення однієї молекули
пальмітинової кислоти становить 96 + 35 –1
= 130 молекул АТФ. + 35 –1
= 130 молекул АТФ.
Поскільки при окисленні ВЖК перетворень зазнає 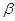 –вуглецевий атом кислот-ного ланцюга, то процес отримав назву –вуглецевий атом кислот-ного ланцюга, то процес отримав назву 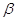 –окислення вищих жирних
кислот. В основному проходить в клітинах печінки. –окислення вищих жирних
кислот. В основному проходить в клітинах печінки.
Активне окислення жирів приводить до накопичення ацетил–КоА (кількість
утвореного ацетил–КоА значно більша ніж можливість його окислення в циклі
Кребса). Надлишкові кількості ацетил–КоА вступають між собою у різноманітні
взаємодії, що приводить до утворення кетонових тіл (ацетооцтова, 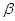 –гідрокси-масляна кислоти,
ацетон, вихідні форми для синтезу холестерину). –гідрокси-масляна кислоти,
ацетон, вихідні форми для синтезу холестерину).
4. Обмін кетонових тіл
       O O
    CH3–C=O + CH3–C=O CH3–C–CH2–C=O + HS–KoA CH3–C=O + CH3–C=O CH3–C–CH2–C=O + HS–KoA
  S–KoA S–KoA S–KoA S–KoA S–KoA S–KoA
ацетил–КоА ацетил–КоА ацетоацетил–КоА
–H2O
–HS–KoA
O
  CH3–C–CH2–C=O CH3–C–CH2–C=O
OH
(ацетооцтова кислота)
–CO2 НАДНН НАД+
CH3–C–CH3 CH3–CH–CH2–COOH
O OH
ацетон  –гідроксимасляна кислота –гідроксимасляна кислота
Ацетооцтова і 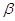 –гідроксимасляна
кислоти–нормальні проміжні продукти обміну ліпідів. Окислюючись в міокарді та
скелетних м‘язах являються важливими енер-гетичними
субстратами. –гідроксимасляна
кислоти–нормальні проміжні продукти обміну ліпідів. Окислюючись в міокарді та
скелетних м‘язах являються важливими енер-гетичними
субстратами.
Ацетон– проміжний продукт при патологічному обміні ліпідів (голодування,
цукровий діабет). Підвищений рівень кетонових тіл у крові приводить до аци-дозу (зміщення активної реакції у «кислу сторону»).
Ацетил–КоА є також вихідною речовиною для синтезу ізопреноїдів, а значить
і стероїдів. Біосинтез стеринового скелету поділяється на три етапи: синтез
мева-лонової кислоти; синтез сквалену; перетворення
сквалену в холестерин.
1.Синтез мевалонової кислоти.
Дві молекули активованої оцтової кислоти (аце-тил–КоА) об‘єднуються утворюючи
ацетоацетил–КоА:
  O O
 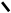 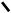 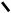 CH3–C=O + CH3–C=O CH3–C–CH2–C=O +
HS–KoA CH3–C=O + CH3–C=O CH3–C–CH2–C=O +
HS–KoA
S–KoA S–KoA S–KoA
ацетил–КоА ацетил–КоА ацетоацетил–КоА
Розгалудження ланцюга відбувається при конденсації молекули
ацетоацетил–КоА із слідуючою молекулою ацетил–КоА:
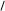 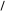     О
H3C OH O О
H3C OH O
        CH3–C–CH2–C=O
+ CH3–C=O –HS–KoA HOOC C C CH3–C–CH2–C=O
+ CH3–C=O –HS–KoA HOOC C C
S–KoA S–KoA CH2
CH2 S–KoA
Утворена 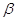 –окси– –окси–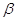 –метилглутарова
кислота в умовах відновлення двома моле-кулами НАДНН відщеплює HS–KoA, в результаті чого карбонільна група пере-творюється в спиртову з утворенням мевалонової кислоти, яка є ключовою спо-лукою для синтезу
ізопреноїдів: –метилглутарова
кислота в умовах відновлення двома моле-кулами НАДНН відщеплює HS–KoA, в результаті чого карбонільна група пере-творюється в спиртову з утворенням мевалонової кислоти, яка є ключовою спо-лукою для синтезу
ізопреноїдів:
      H3C OH
O H3C OH H3C OH
O H3C OH
    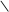 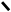  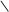   HOOC C C 2НАДНН 2НАД HOOC C CH2 HOOC C C 2НАДНН 2НАД HOOC C CH2
 CH2 CH2 S–KoA –НS–KoA CH2 CH2 OH CH2 CH2 S–KoA –НS–KoA CH2 CH2 OH
3,5-диоксі-3-метилвалеріанова кислота
(мевалонова кислота)
2.Утворення активного ізопрену та його перетворення в сквален.
Послідовне фосфорилювання мевалонової кислоти молекулами АТФ приводить до
утво-рення «активного ізопрену» (ізопентенілпірофосфату). При конденсації
ізопенте-нілпірофосфату з 3,3–диметилалілпірофосфатом утворюється
геранілпірофосфат, дві молекули якого утворюють сквален шляхом відновлювальної
конденсації в присутності НАД-залежних дегідрогеназних систем. Дана стадія
протікає в роз-чині цитоплазми за відсутності кисню:
    H3C OH
H3C OH H3C OH
H3C OH
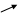       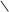  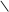   HOOC C CH2 АТФ АДФ HOOC C CH2 HOOC C CH2 АТФ АДФ HOOC C CH2
 CH2 CH2 OH CH2
CH2 OФ CH2 CH2 OH CH2
CH2 OФ
5-фосфомевалонова
кислота
    H3C OH
H3C OH H3C OH
H3C OH
 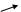      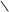  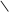   HOOC C CH2 АТФ АДФ HOOC C CH2 HOOC C CH2 АТФ АДФ HOOC C CH2
 CH2 CH2 OФ CH2
CH2 O–Ф–Ф CH2 CH2 OФ CH2
CH2 O–Ф–Ф
5-пірофосфомевалонова кислота
   H3C OH
H3C H3C OH
H3C
  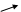       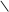   HOOC C CH2 АТФ АДФ C CH2 HOOC C CH2 АТФ АДФ C CH2
 CH2 CH2 OФ–Ф –СО2
–Н2О CH2 CH2 O–Ф–Ф CH2 CH2 OФ–Ф –СО2
–Н2О CH2 CH2 O–Ф–Ф
ізопентенілпірофосфат
  H3C
H3C H3C
H3C
        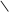  C CH2 C CH2 C CH2 C CH2
 CH2 CH2 OФ–Ф CH3
CH O–Ф–Ф CH2 CH2 OФ–Ф CH3
CH O–Ф–Ф
ізопентенілпірофосфат 3,3-диметилалілпірофосфат
  H3C H3C H3C H3C
         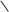  C CH2 + C CH2 –Н4Р2О7 C CH2 + C CH2 –Н4Р2О7
CH2 CH2 OФ–Ф CH3 CH O–Ф–Ф
ізопентенілпірофосфат 3,3-диметилалілпірофосфат
3. Перетворення сквалену в холестерин. На стадії
окислення відбувається цик-лізація і утворюються
ланостерин і холестерин. При цьому відбувається наси-чення
подвійних зв‘язків і окислювальне відщеплення метильних груп при четвертому і
чотирнадцятому атомах вуглецю у вигляді СО2. Циклізація сквалену в
структуру що нагадує стероїди, як і послідуючі реакції, протікає на поверхні
ендоплазматичного ретикулуму:
Біосинтез стероїдних гормонів
Вихідною сполукою в біосинтезі стероїдних гормонів є холестерин. Із
ньо-го послідовно утворюються прегненолон і прогестерон. Прогестерон –
ключова сполука в синтезі всіх стероїдних гормонів. Сам по собі володіє значною
біоло-гічною активністю як гормон жовтого тіла (гестаген).
Прегнандіол, що виділя-ється із сечею,– неактивний продукт
розпаду прогестерону.
Біосинтез стероїдних гормонів, що починається з прогестерону, каталізу-ється стереоспецифічними ферментами (гідроксилазами). В назву
гідроксилази входить номер який вказує номер вуглецевого атома по якому
відбувається при-єднання гідроксильної групи.
Відщеплення бокового ланцюга у молекулі прогестерону приводить до
утворення андростендіону, при відновленні якого утворюється тестостерон.
Тестостерон і його аналоги володіють анаболічною активністю. Використанню
цих сполук в якості лікувальних анаболічних препаратів зашкоджує їх чітко
ви-ражена андрогенна дія.
В зв‘язку з цим було синтезовано нові стероїдні сполуки, близькі по
структурі до андрогенів, що володіють вибірковою анаболічною активністю при
маловира-женій андрогенній дії. Вони отримали назву «анаболічні стероїди».
Найбільш характерною властивістю анаболічних стероїдів є їх здатність до
сти-мулювання синтезу білка в організмі. Вони позитивно впливають на азотистий
обмін; викликають затримку азоту в організмі, зменшують його виділення через
нирки із сечею. Також відбувається затримка виділення необхідних для синтезу
білка калію, сірки і фосфору. Анаболічні препарати сприяють також фіксації
кальцію у кістках. Клінічна дія анаболічних стероїдів проявляється у підвищенні
апетиту, збільшенні маси тіла, покращенні загального стану хворих, більш
швид-кій кальцинації кісток(при остеопорозі).
Препарати, що відносяться до групи анаболічних стероїдів, володіють різ-ним по силі і тривалості анаболічним ефектом. Вони також
відрізняються по від-носній андрогенній активності.
МЕТАНДРОСТЕНОЛОЛ (METHANDROSTENOLOLUM).
Синоніми: Діанабол, Неробол, Anabolex, Anabolin, Anaboral, Bionabol, Danabol, Distranorm,
Lanabolin, Metanabol, Metandienone, Nabolin, Nerobol, Novabol, Perabol,
Vanabol.
Володіє андрогенною активністю, проте менш активний тестостерону.
Викликає виражений анаболічний ефект. Андрогенна дія метандростенолону у 100
раз менша аналогічної дії тестостерону при майже однаковій анаболічній
активності. Основними критеріями до прийому метандростенололу, як і інших
анаболічних стероїдів, є : порушення білкового обміну, астенії, інфекційні та
інші захворю-вання що пов‘язані із втратою білка. Препарат призначають після
важких травм, операцій та опіків.
В ендокринологічній практиці препарат та його аналоги застосовують при:
гіпо-фізарній та наднирниковій недостатньості, токсичному зобі, стероїдному
діабеті, гіпофізарній карликовості.
В терапевтичній практиці назначають при хронічній коронарній
недостатньості та інфаркті міокарду, міокардитах, ревматичних ураженнях серця
та атероскле-ротичному кардіосклерозі; при виразковій
хворобі шлунку та дванадцятипалої кишки, хронічних захворюваннях нирок
(супроводжуються втратою білка) та ле-гень. Ці препарати
використовуються також для лікування остеопорозу, великих переломів, при
міопатіях і прогресуючій м‘язевій дистрофії.
Лікування метандростенолоном повинно відбуватися під керівництвом лі-каря. При прийомі препарату можливі слідуючі побічні явища: диспептичні
роз-лади, збільшення печінки, виникнення жовтухи та
набряків. У жінок (особливо при тривалому прийомі) можливо порушення
менструального циклу, огрубіння голосу, посилення росту волосся по чоловічому
типу.
Тривале безперервне приймання стероїдних препаратів приводить до ус-кладнень та порушення процесів обміну кальцію, гормонального
фону організ-му, структурних змін у тканинах нирок,
серцевого м‘язу, печінки.
До групи анаболічних стероїдів також відносяться:
ФЕНОБОЛІН (PHENOBOLINUM)
Синоніми: Дураболін, Нандролонфенілпропіонат, Нероболіл, Тураболіл, Туріна-бол.
Показання до застосування такі як і для попереднього препарату, особливо
у ви-падках коли необхідно досягти сильнішої анаболічної і
антикатаболітичної дії.
РЕТАБОЛІЛ (RETABOLIL).
Препарат протипоказаний при онкозахворюваннях молочної та передміхурової
залоз, вагітності. Необхідна обережність при недостатній функції нирок та
печінки.
СІЛАБОЛІЛ (SILABOLINUM)
Сілаболіл володіє пролонгованою анаболічною дією. Після одноразового вве-дення ефект триває 10–14 днів. Володіє незначною андрогенною
дією.
5. Біосинтез гліцерину
Вихідною речовиною для даного процесу є фосфодиоксіацетон, який віднов-люється НАД-залежними дегідрогеназами до фосфогліцеринової
кислоти. Остання під дією ферменту гліцерофосфатази розщеплюється з утворенням
глі-церину і фосфорної кислоти:
    CH2–OH СН2–ОН CH2–OH СН2–ОН
     C=O OH НАДНН НАД+ СН–ОН
ОНгліцерофосфатаза C=O OH НАДНН НАД+ СН–ОН
ОНгліцерофосфатаза
 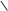 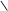 CH2–O–P=O СН2–О–Р=О CH2–O–P=O СН2–О–Р=О
OH ОН
 СН2–ОН СН2–ОН
 СН–ОН + Н3РО4 СН–ОН + Н3РО4
СН2–ОН
6. Біосинтез вищих жирних кислот
Проходить за участю двох типів ферментних систем: мітохондріальної і
цитоплазматичної.
Мітохондріальна – забезпечує подовження вуглець-вуглецевого
ланцюга вже існуючої кислоти шляхом приєднання молекули ацетил–КоА.
Цитоплазматична – забезпечує синтез нової молекули кислоти з
молекули аце-тил–КоА. Для її роботи необхідним є наявність
НАДФ та біотин–залежних фер-ментних систем, синтетази
жирних кислот, АТФ, гідрокарбонатних аніонів.
Процес умовно поділяють на підготовчу і основну стадії.
Підготовча стадія полягає в активації молекули вуглекислого газу
(СО2), і її вза-ємодії з молекулою ацетил–КоА з
утворенням активної форми малонової кислоти (малоніл–КоА). Закінчується
взаємодією щойноутвореної малонової кислоти з новою молекулою ацетил–КоА з
утворенням фермент–субстратного комплексу. Дану стадію регулює ферментна
система – синтетаза вищих жирних кислот:
Основна стадія – полягає в перетворені фермент-субстратного комплексу у
фрагмент жирної кислоти:
           CH3 СH3 CH3 СH3 CH3 СH3 CH3 СH3
  C=O НАДНН НАД+ CH–OH –H2O CH НАДНН НАД+
CH2 C=O НАДНН НАД+ CH–OH –H2O CH НАДНН НАД+
CH2
 CH2 CH2 CH CH2 CH2 CH2 CH CH2
     C=O HS C=O
HS C=O HS C=O HS C=O HS C=O
HS C=O HS C=O HS
   S–Ф
S–Ф S–Ф S–Ф S–Ф
S–Ф S–Ф S–Ф
Новоутворений фрагмент переноситься на переферичну тіолову групу, а до
центральної приєднується нова молекула малоніл –КоА, утворюючи новий фер-мент-субстратний комплекс:
CH3 CH3 COOH CH3  CH2 CH2
CH2 + CH2 CH2  CH2 CH2
CH2 C=O CH2      C=O HS C=O HS
C=O S–KoA C=O
   S–Ф S–Ф S S–Ф S–Ф S
             HS –HS–KoA COOH HS –HS–KoA COOH
 CH2 Ф CH2 Ф
 C=O C=O
S
Приєднання фрагментів малоніл–КоА відбувається до
тих пір, поки молекула жирної кислоти не набере необхідну
кількість вуглецевих атомів. Після цього
фермент-субстратний комплекс розщеплюється на власне фермент і активну форму
кислоти:
      CH3 CH3 CH3 CH3
     CH2 CH2 HS CH2 CH2 HS
 CH2 7 CH2 7 + Ф CH2 7 CH2 7 + Ф
   C=O C=O HS C=O C=O HS
 S S
 HS HS
пальмітил–ферментний комплекс пальмітил–КоА ферментний комплекс
Отже: ацетил–КоА – основна вихідна речовина а малоніл–КоА –
основний мета-боліт процесу синтезу вищих жирних кислот.
Для утворення молекули пальміти-нової кислоти
ферментативні реакції основної стадії повторюються 7 раз, а для утворення
стеаринової кислоти – вісім разів. Одноненасичені кислоти синтезу-ються
з відповідних насичених кислот, а поліненасичені (лінолева, ліноленова,
арахідонова) в організмі не синтезуються.
7. Біосинтез тригліцеридів
Трьохстадійний процес. Вихідною речовиною для синтезу є похідне гліцерину
– гліцеринфосфорна кислота та активовані форми вищих жирних кислот.
1. Утворення фосфатидної кислоти:
            CH2–OH CH2–O–C(O)–C17H35 CH2–OH CH2–O–C(O)–C17H35
CH–OH + 2 C17H35COSKoA –HS–KoA CH–O–C(O)–C17H35
        CH2–O–P=O CH2–O–P=O CH2–O–P=O CH2–O–P=O
HO OH HO
OH
2. Ферментативне розщеплення фосфатидної кислоти за
допомогою фосфо-ліпази С:
CH2–O–C(O)–C17H35 CH2–O–C(O)–C17H35
CH–O–C(O)–C17H35 H–OH CH–O–C(O)–C17H35
+ H3PO4
CH2–O–P=O CH2–OH
HO OH
3.Утворення тригліцериду:
 CH2–O–C(O)–C17H35 O=C–C17H35 CH2–O–C(O)–C17H35 CH2–O–C(O)–C17H35 O=C–C17H35 CH2–O–C(O)–C17H35
 CH–O–C(O)–C17H35
+ KoAS CH–O–C(O)–C17H35 CH–O–C(O)–C17H35
+ KoAS CH–O–C(O)–C17H35
CH2–OH –HS–KoA CH2–O–C(O)–C17H35
Поскільки фосфатиди і нейтральні жири мають подібну будову, то деякі етапи синтезу для них подібні. Спільним
метаболітом для обох процесів є 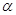 , ,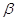 –ди-гліцериди. –ди-гліцериди.
8. Біосинтез фосфогліцеридів
1.Фосфорилювання азотистої основи:
 OH OH
   (CH3)3–N–CH2–CH2–OН АТФ АДФ (CH3)3–N–CH2–CH2–O–P=O (CH3)3–N–CH2–CH2–OН АТФ АДФ (CH3)3–N–CH2–CH2–O–P=O
OH
2.Активування фосфохоліну з утворенням
цитидиндифосфохоліну:
               OH О
О OH О
О
   (CH3)3–N–CH2–CH2–O–P=O ЦТФ Н4Р2О7
(CH3)3–N–CH2–CH2-O-P-O-Р-О-Ц (CH3)3–N–CH2–CH2–O–P=O ЦТФ Н4Р2О7
(CH3)3–N–CH2–CH2-O-P-O-Р-О-Ц
OH ОН ОН
3.Утворення фосфатиду:
CH2–O–C(O)–C17H35 O
O
 CH–O–C(O)–C17H35 + (CH3)3–N–CH2–CH2-O-P-O-Р-О-Ц CH–O–C(O)–C17H35 + (CH3)3–N–CH2–CH2-O-P-O-Р-О-Ц
CH2–OH OH
OH –ЦМФ
  CH2–O–C(O)–C17H35 CH2–O–C(O)–C17H35
 CH–O–C(O)–C17H35 CH–O–C(O)–C17H35
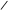 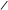  CH2–O–P–O–CH2–CH2–N(CH3)3 CH2–O–P–O–CH2–CH2–N(CH3)3
O OH
Питання для самоконтролю
1.Ферментативне перетворення ліпідів у травному тракті людини.
2.Роль жовчі у процесах розщеплення ліпідів.
3.Транспортні форми ліпідів та шляхи розповсюдження по організмі.
4.Окислення гліцерину. Енергетичний ефект процесу.
5. Окислення вищих жирних кислот. енергетичний ефект процесу.
6. Обмін кетонових тіл. Нормальні та аномальні продукти
обміну.
7. Синтез холестерину та стероїдних гормонів. Біологічна роль
стероїдних гор-монів.
8. Синтез гліцерину.
9. Синтез вищих жирних кислот.
10. Синтез фосфогліцеридів.
11. Біологічна роль сполук класу ліпіди.
|