Курсовая работа: Возможности и перспективы использования ильменей типичных водоемов аридной зоны для создания озерных рыбоводных хозяйств дельты реки Волги
Курсовая работа: Возможности и перспективы использования ильменей типичных водоемов аридной зоны для создания озерных рыбоводных хозяйств дельты реки Волги
Введение
Волго-Каспийский бассейн
исторически является житницей России, поставляя в отдельные периоды развития
страны до 80 % добываемой ею рыбы. Северный Каспий и дельта Волги – район
уязвимый в экологическом отношении. Это обусловлено, с одной стороны, высокой
антропогенной активностью в бассейне моря, с другой – неустойчивостью
гидрологического режима реки, моря и происходящих в них биопродукционных
процессов.
В настоящее время сток
реки Волги практически полностью зарегулирован Волго-Камским каскадом
водохранилищ. В бассейне реки Волги проживает более четверти населения России и
производится свыше 20 % промышленной продукции.
Однако, рыбоводство
развивается значительно медленнее и даже на современном этапе находится лишь на
стадии поиска оптимальных путей увеличения биопродуктивности, использования
водоемов. Это объясняется многими причинами, среди которых важнейшей является
возможность удовлетворения спроса в рыбной продукции за счет естественных
водоемов. Рыбу проще ловить, и до прошлого века существовала уверенность в том,
что запасы рыбы неисчерпаемы, для бесконечного увеличения, промысла необходимо
лишь совершенствовать орудия промысла. Но численность рыб в естественных
водоемах стала неуклонно снижаться. На данный момент процесс нарастает большими
темпами, что явилось причиной все большего возрастающего интереса к искусственному
разведению рыбы.
Астраханская область
имеет большие возможности увеличения рыбы из естественных водоемов. В настоящее
время основными местами рыболовства являются река Волга и Каспийское море.
Добыча реки в выше
указанных водоемах весьма интенсивна и ее дальнейший рост в ближайшей
перспективе маловероятен. Однако, Волгой и Каспийском морем не ограничен весь
перечень водоемов, где возможен эффект рыболовства, одним из таких резервов
являются Западно – подстепные ильмени. Площадь этих ильменей ранее составляла
около 600 тыс. га
Из этой площади под
выростные и нагульные пруды освоено около 1 % имеющегося фонда. В связи с
наметившимся в последнее время дефицитом пресной воды перспективным
представляется использование высоко минерализованных водоемов для интенсивного
рыборазведения.
Целью данной работы
является определение возможности и перспективы использования ильменей типичных
водоемов аридной зоны для создания озерных рыбоводных хозяйств дельты реки
Волги.
Задачами данной дипломной
работы является:
1. Исследование качественного состава
населения ильменных водоемов.
2. Исследование количественного состава
населения ильменных водоемов.
3. Изучение годовой динамики планктонных
и бентосных организмов за период с 1996 по 2003 гг.
4. Обоснование возможностей
использования ильменных водоемов в рыбохозяйственных целях.
Глава1. Литературный обзор
1.1.
Характеристика малых водоемов комплексного назначения
Малые водоёмы – пруды,
водохранилища, озёра и другие площадью до 1 тыс.га, на которые не
распространяются Правила об охране рыбных запасов и регулирования рыболовства в
водоёмах, использующихся для ирригации, рыборазведения, водопоя скота и других
целей, получили название водоёмов комплексного назначения (ВКН). Эти водоёмы, в
основном, находятся в землепользовании сельскохозяйственных предприятий.
Многие сельхозпредприятия
используют водные площади для увеличения производства пищевой продукции и
осваивают ВКН для выращивания рыбы на ВКН возможно без применения искусственных
кормов (зерно, комбикорма и т.п.) на естественной пищевой базе, поэтому
рыбопродуктивность в среднем составляет 2-4 ц/га.
Учитывая тот факт, что по
гидрологическим, химическим и биологическим параметрам приспособленные водоёмы
комплексного назначения коренным образом отличаются от классических нагульных
рыбоводных прудов, необходимо применять на них специальную технологию
рыбоводства.
Условно по способу
накопления и возможности сброса воды ВКН подразделяются на четыре категории.
Овражно-балочные
запрудные;
карьерно - котловинные
наливные;
поименно-лагунные
мелководные;
русловые проточные.
Одновременно водоёмы в
связи с особенностью их рыбохозяйственного освоения по размерам разделены на
три группы: малые – до 5 га, средние – от 50 до 300 га и крупные – от 300 до
1000 га.
Овражно-балочные запрудные
водоёмы наполняются талыми или дождевыми водами, имеют одну плотину, у которой
отмечается максимальная глубина. Благодаря естественному перепаду уровней – от
2-3 до 8-10 м – имеется возможность полного сброса воды и рыбы через
рыбоуловитель. В таких водоемах наблюдается слоистость вод по температуре и
содержанию кислорода у дна и поверхности. Кормовая база в горных и предгорных
зонах незначительна, в равнинных – может быть высокая. Минерализация в зонах
ирригации превышает норму, обычно принятую для нагульных карповых прудов, что
предполагает выращивание и солелюбивых рыб. Эта категория водоёмов наиболее
перспективная в освоении, так как не требует затрат на мелиорацию ложа и
организацию промысла. Площади обычно до 50 га, реже до 300 га. Среди рыб – естественных
обитателей водоёмов – доминируют мелкие формы: пескари, плотва и другие,
моллюски редки, в горных водоёмах нет личинок хирономид. Рыбопродуктивность в
горных и пустынных зонах до 2-4 ц/га, в других регионах до 6-8 ц/га.
Карьерно - котловинные наливные
водоёмы наполняются, как правило, подпочвенными водами или за счёт водоподачи
по каналам и водоводам. Могут быть созданы в торфяниках, каменистых и песчаных
карьерах, плотины, как правило, не имеют. Максимальная глубина водоёма от 8-10
до 15 метров может быть в любом месте; берега обрывистые, на ложе могут быть
отдельные ямы. Естественного стока воды нет. Слоистость вод по температуре –
обязательное явление благодаря подземным источникам. Прогревание нижних слоёв
из-за отсутствия прочности очень медленное, чаще отмечается постоянная
температура у дна 8-10 . В торфяных карьерах встречаются сплавины, корневища
деревьев. Вода в торфяных карьерах имеет, кислую среду (рН ниже 7), что
сдерживает возможность нереста карповых рыб. В каменисто-песчаных водоёмах, как
правило, рН больше 7. Кормовая база постоянно низкая. В торфяных карьерах
развитию зоопланктона реже предшествует интенсивное развитие фитопланктона.
Среди рыб доминируют линь, золотой карась, ротан, реже щука. В
каменисто-песчаных – пескарь, окунь, щука, сом, серебряный карась. Вылов рыбы
возможен неводами на подготовленных тонях, куда она привлекается подкормкой, а
также с применением ставных неводов, верш, электролова. Лучше облавливаются
карьеры после предварительной частичной откачки воды. Площадь торфяных карьеров
– до 50-300 га, каменисто-песчаных чаще до 50, реже – до 300 га.
Рыбопродуктивность – до 2-3 ц/га.
Пойменно-лагунные
мелководные водоёмы отличаются ровным ложем и относительным мелководьем. К этой
категории могут быть отнесены водоёмы лиманного типа, построенные на поймах и
других естественных понижениях суши. Наполняются водоёмы в половодье
(пойменные), при соединении их с морем (лагунные) или затоплением морской или
пресной водой естественных погруженных частей суши (лиманы). Известно также
заполнение механическое за счет ирригационных сборных и артезианских вод
различных понижений обширной площади. Плотины, если они есть, низконапорные.
Максимальная глубина 2-3 м, ложе пологое, ровное. Возможность естественного
стока, кроме водоёмов, размещённых выше окружающего ландшафта, отсутствует.
Прогреваемость до дна, проточность могут отсутствовать. По качеству воды
выделяются пресные водоёмы и с водой различной солёности, вплоть до океанской –
32-35%о и выше, пригодные для выращивания солелюбивых рыб. Среди рыб в резко
осолоненных водоёмах превалируют 3-5 видов рыб – атерина, колюшки, реже кефаль.
В пресноводных – большой состав лиманной ихтиофауны – плотва, краснопёрка,
щука, окунь, линь, бычок и другие. Кормовая база может быть значительная, обеспечивающая
получение 8-10 ц/га рыбы. Из кормовых организмов превалируют нектобентические
формы – гаммариды, мизиды, а также черви и личинки хирономид. Зарастаемость
может быть полная, как погруженной мягкой – рдесты, уруть, хара, так и
тростником, рогозом и другой жесткой растительностью. “Цветение” воды в этих
водоёмах – обычное явление.
В рыболовстве такие
водоёмы используются путем вселения карпа, толстолобика, сома в пресноводные,
кефалей, полосатого окуня, осетровых – в солоноватоводные и активного промысла
неводами с практически полным отловом вселенцев. Площадь этих водоёмов от
50-300 га до 1 тыс. га и более. Рыбопродуктивность 6-8 до 10 ц/га.
Русловые проточные
водоемы, построенные на речках и малых реках, наполняются водой постоянно.
Создаются за счёт подпора реки в удобном по ландшафту месте. Максимальная
глубина у плотины и у затопленного русла. Берега могут быть пологими и
обрывистыми, но как правило, хотя бы одно пологое место в верховьях водоёма
имеется. Возможности полного или даже частичного сброса воды нет. Слоистость
вод по температуре и кислороду благодаря постоянному водообмену отсутствует.
Прогреваемость практически равномерная, чаще наиболее прогреты слои воды у
плотины. Кормовая база относительно выше у плотины, но постоянное присутствие
местных рыб (окунь, щука, плотва, пескари, караси и т.д.) способствует высокой
конкуренции с разводимыми рыбами. Качество воды чаще высокое, что
обеспечивается постоянной её сменой. В рыбоводстве могут быть использованы при
наличии рыбозащитных устройств на водоподаче и сбросе. Рыбопродуктивность от
2-4 до 6 ц/га.
Режим эксплуатации
водоёма при выращивании рыбы связан с особенностями наполнения и сброса воды.
Овражно-балочные водоёмы, как правило, однолетнего заполнения. При выращивании
теплолюбивых рыб: карпа, толстолобика, амура и других период эксплуатации
связан со сроками наполнения водой весной и прекращением роста рыбы осенью. В
случае выращивания сиговых и других холодолюбивых рыб период выращивания может
быть продлён и в зимнее время.
Карьерно-котловинные
наливные водоёмы – многолетнего регулирования. Режим выращивания рыбы,
плотность посадки ее связаны с продолжительностью ледостава и газовым режимом.
В торфяных карьерах летние заморы – частое явление, поэтому рыбы, требующие
высокого содержания кислорода (осетровые, лососевые, сомы и другие), для таких
категорий водоёмов не пригодны.
Пойменно-лагунные и
другие мелководные водоёмы, используемые для полива, могут иметь значительное
уменьшение площади. Зарыбление таких водоёмов рассчитывается на 70% площади при
начале сработки уровня с июля-августа, из расчета 50% площади при начале
сработки уровня в весеннее время. При обеспечении глубины отдельных участков
3-4 м, где рыба может переносить зимовку, рекомендуется осеннее зарыбление.
Русловые водоёмы имеют
максимальный уровень в паводковый период. При отсутствии надежных рыбозащитных
устройств (РЗУ) зарыбление проводится после прохождения паводка. В случае
надежности РЗУ рекомендуется осеннее зарыбление.
ВНК должны быть
обеспечены водой, не загрязненной ядохимикатами, применяемыми в сельском
хозяйстве, и сточными водами промышленных сельскохозяйственных и других
предприятий, не имеющих необходимый газовый и термический режим, отвечающий
требованиям ОСТ 15.372.87.
Вокруг водоема
устанавливают водоохранную зону шириной 500 м от границы затопления при
максимальном стоянии паводковых вод или 2000 м от берегов в период между
паводками.
В водоохраной зоне (не
менее 2000 м от существующих берегов) запрещается:
- строительство складов
для хранения пестицидов, взлетно-посадочных площадок для самолетов и
вертолетов;
- размещение
животноводческих построек, силосных ям и ванн для ветеринарно-санитарных
обработок животных;
- устройство площадок для
протравления семян и приготовления отравленных приманок, рабочих растворов
ядохимикатов и заправка ими машин;
- устройство мест
обезвреживания техники и тары из-под пестицидов.
Запрещается производить
авиаопыливание, авиаопрыскивание и аэрозольные обработки посевов ближе 2000 м
от существующих берегов рыбохозяйственных водоемов.
Для предотвращения
попадания в ВКН загрязненного поверхностного стока санитарную зону засевают
лесными полосами, кустарниками и травами. В ВКН запрещается мытье спецодежды,
тары, техники, используемой для применения вредных химических веществ.
ВКН заселяются
рыбопосадочным материалом из рыбопитомников ( желательно постоянно за одного и
того же ), благополучного по заразным болезням рыб. Перевозка рыбы разрешается
только при наличии ветеринарного свидетельства (форма 1), где должно быть
указано, что хозяйство благополучно по инфекционным и инвазионным болезням рыб,
и рыба подвергнута профилактической противопаразитной обработке. Перевозить и
пересаживать рыбу следует с соблюдением мер предосторожности, не допуская
травмирования; в пути контролировать содержание кислорода и поддерживать
существующую температуру воды.
1.2.
Организация и использование пастбищных озерных хозяйств в солоноватоводных
водоемах Астраханской области
В дельте Волги, площадь
которой составляет 21 тыс. км2., имеется около 600 тыс. га.
Озеровидных водоемов- ильменей, в том числе 300 тыс. га в ее западной
подстепной части. Эти мелководные, хорошо прогреваемое водоемы имеют постоянную
или временную связь с Волгой. Большинство из них имеют рыбохозяйственное значение:
здесь находятся места (размножения и нагула рыб осуществляется промысловый лов
рыбы. Озера, полностью потерявшие связь с рекой, обычно сильно засолены и не
имеют рыбохозяственного значения. Основу рыбного населения озер ильменей
составляют красноперка, окунь, густера, карась, линь, щука. Встречаются также
вобла, сазан, сом, лешь, язь, берш и некоторые другие виды рыб. Уловы рыбы в
озерах дельты до недавнего времени, по оценочным данным составляли около 12
тыс. т/год. Однако, в последний годы, в связи с ухудшением условий обитания рыб
под воздействием антропогенных факторов, они заметно упали, и в 1994 г.
составили около 5 тыс. т млн. в среднем всего 9 кг рыбы с каждого гектара.
Такие уловы не соответствуют потенциально возможным. (Сальников, Киселева.
1994).
Рыбохозяйственный фонд
озер дельты Волги используется неудовлетворительно. У озер оказалось слишком
много "хозяев", интересы которых часто совершенно не связаны с рыбным
хозяйством. Озерами владеют местные районные администрации, управления
оросительных систем, отдельные сельскохозяйственные колхозы и совхозы,
выступающие сейчас под другими названиями, различные товарищества с
ограниченной ответственностью и др. На озерах процветает браконьерский лов.
Многие фермеры, взявшие озера в аренду для рыбохозяйственных целей, рыбоводные
работы в полном объеме не осуществляют, занимаются преимущественно не
выращиванием рыбы, а только ее ловом и продажей. Органы рыбоохраны и одного хозяйства
(Госкомвод) не всегда обеспечивают должный контроль за соблюдением на озерах
рыбохозяйственного и водохозяйственного законодательств не пресекают действия отдельных
водопользователей, нарушающих естественный режим озер. В настоящее время в
дельте Волги в целом сложились неблагоприятные условия рыболовства: резко упали
уловы осетровых, крупного и мелкого частика, что связано не только с состоянием
их запасов, но и с изменением условий промысла в результате подъема уровня
Каспия. Оказались подтопленными многие тоневые участки, усложнились условия
лова рыбы ловушками (секретами) и т. д. Не лучше сложилось положение и с
прудовым товарным рыбоводством. В 1994 г. прудовой рыбы было получено всего
около 3.9 тыс. т. В настоящее время рыбная продуктивность прудов упала до 530
кг/га. Причин этому множество: дороговизна кормов и удобрений, но главное, вероятно,
в том, что полностью потеряно управление этой отраслью рыбного хозяйства. В
сложившихся условиях, для увеличения уловов рыбы в дельте Волги, видимо,
необходимо обратить самое серьезное внимание на создание на базе озер- ильменей
дельты пастбищных нагульных рыбоводных хозяйств с преимущественным выращиванием
на естественной кормовой базе растительноядных рыб китайского фаунистического
комплекса (рыб низкого трофического уровня) белого и пестрого толстолобиков, их
гибрида и белого амура. Выращивать этих рыб надо в поликультуре с карпом и
осетровыми, которые нуждаются в кормлении искусственными кормами. В качество
добавочной рыбы можно использовать хищника (щука или судак). Из
интенсификационных мероприятий в озерах предусматривается лишь внесение удобрений.
При этом в зависимости естественной биологической продуктивности и кормности
отдельных озер может быть получено товарной рыбы от 400 до 2000 кг/га/год, а
при кормлении еще выше. Организация пастбищных озерных рыбных хозяйств
потребует увеличения производства рыбопосадочного материала, расширения сети
рыбопитомников и повышения эффективности их работы (Сальников, Киселева. 1994).
При полном освоении
пригодных для организации пастбищного рыбоводства озер-ильменей дельты Волги,
уловы рыбы в дельте в обозримой перспективе могут возрасти в 1,5-2 раза, по
сравнению с современными. Озерное рыбоводство, вероятно, должно стать
приоритетным направлением развития аквакультуры в устьевой области реки Волги.
Для этого необходимо разработать специальную региональную программу, которая
могла бы войти составной частью Государственной федеральной программы развития
рыбного хозяйства.
В зоне западно-подстепных
ильменей расположены два хозяйства, входящие в состав ООО
«Астраханьрыбагрогаз», — РП «Камышовское» и СХП «Озерный». И хотя суммарный
фонд ильменей насчитывает 60 единиц, общей площадью более 8 тыс. га, в силу
различных причин в рыбоводстве используется только 19 ильменей общей площадью
2,6 тыс. га. На такой площади при грамотном ведении хозяйства с применением органических
и минеральных удобрений без затрат на кормление ежегодно можно получать до
1,3-1,5 тыс. т. рыбной продукции. При кормлении рыбы измельченной зеленой
массой (люцерна, тростник и др.) можно увеличить рыбопродуктивность имеющейся
прудовой площади до 1,8-2,0 тыс.т в год. (Меретин,2002).
В 2002 г. вступил в строй
СХП «Икрянинский рыбозавод». Согласно программе развития рыбной отрасли ООО
«Астраханьрыбагрогаз» на 2000-2005 гг. в 2001 г. завод переработал 5000 т
рыбного сырья от собственного вылова, выращивания и закупки ( Меретин,2002).
До 2005 г. будут
постепенно наращиваться мощности Икрянинского рыбозавода и увеличиваться объемы
сырья до 14 тыс. т в год (собственный вылов — б тыс. т, закупка — 5 тыс. т,
прудовое рыбоводство — 3 тыс. т).
Увеличение производства
прудовой рыбы до 4,0 тыс. т. в год реально можно достичь путем расширения
прудовых (ильменных) площадей с 2,5 тыс. га до 7 тыс. га, одновременно повышая
рыбопродуктивность водоемов за счет повышения интенсификационных мероприятий
(органические и минеральные удобрения, кормление рыбы зеленой массой, выкос
растительности и др.).
Практика показывает, что
средняя стоимость мелиоративных работ по преобразованию ильменей в
озерно-товарное хозяйство составляет 15-17 тыс. руб. за 1 га. За последние два
года (2001-2002) на прудовое рыбоводство выделялось 25 млн. руб.
Подготовка нагульных
водоемов включает в себя строительство водоподающих каналов сбросной системы,
мелиорацию ложа водоемов (удаление высшей растительности, глубокая вспашка,
нарезка сбросных коллекторов) и др. Всего необходимо ввести в рыбохозяйственный
оборот не менее 7 тыс. га ильменей. Потенциал наращивания рыбоводных площадей в
хозяйствах имеется. Наличие воспроизводственного участка позволяет зарыблять
нагульные водоемы в гораздо более ранние сроки, чем при приобретении
рыбопосадочного материала на стороне. (Меретий2002).
Однако, реализация данной
программы тормозится по ряду причин. Во-первых, с каждым годом все более
усложняется процедура отвода и лицензирования водоемов под озерно-товарные
хозяйства, как на районном, так и на областном уровне. Во-вторых, высокая
стоимость мелиоративных работ и обустройства ильменей. Все это негативно
сказывается на темпах развития прудового рыбоводства. (Меретин,2002).
Для нормального
функционирования рыбной отрасли необходимо развивать вспомогательное
производство, включающее мелиоративный отряд, транспортное хозяйство,
энергетическую службу, ремонтные мастерские. (Меретин.2002).
Реализация программы «Развитие
рыбной отрасли ООО Астраханьрыбагрогаз» позволит: рационально эксплуатировать
многочисленные ильмени и межбугровые впадины; получить в больших объемах разнообразную,
в том числе деликатесную, рыбную продукцию и тем самым загрузить мощности
строящегося рыбозавода; реанимировать забытые районы промысла; нормализовать
санитарное состояние водоемов; использовать методы рыбоводной мелиорации для
повышения плодородного возвращения в сельскохозяйственное пользование
потерянных ранее земель; создать дополнительные рабочие места; решить некоторые
социальные вопросы (дороги, питьевой трубопровод в хозяйствах, жилищное
строительство); увеличить поток налоговых отчислений во все уровни бюджета.
(Меретин,2002).
1.3 Общая характеристика
биопродуктивности водоемов Астраханской области
1.3.1
Современное состояние фитопланктона Западных подстепных ильменей
Фитопланктон является
основным источником первичной продукции органического вещества, он часто
выполняет роль главной трофической основы жизни в водоеме. Кроме того,
фитопланктон является чутким индикатором меняющихся условий внешней среды, что
довольно часто отражается на динамике его развития. И, наконец, фитопланктон
часто служит основным пищевым объектом некоторых видов промысловых рыб. Поэтому
в условиях давно развитой рыбной промышленности Волго-Каспийского района именно
здесь в свое время возникли важные идеи у крупнейшего биолога 19 века Карла
Бэра о продуктивности водоемов (Кражан,1971).
С начала прошлого
столетия началось научное исследование водоемов. Одновремённо с исследованием
других компонентов планктона параллельно проводились исследования водорослей.
Так, летом 1907 года в речном русле против Астрахани в первой половине лета до
середины июня было обнаружено 20 видов водорослей, на время после середины июля
пришлось всего 7 видов.
Первый список организмов,
найденных в дельте Волги и ее притоков был опубликован М.Х. Сергеевой в 1909
году, в трудах Астраханской ихтиологической лаборатории. Список состоит из 390
видов: из них -219 растительных форм, 11 - простейших, 31- коловраток, 38
-низших ракообразных (27 ветвистоусых и 11 веслоногих), 35-высших раков, 13-других
беспозвоночных (кишечнополостных, мшанок и прочих) и 43-вида рыб. (Сокольский,
1995).
Круглогодичные наблюдения
над фитопланктоном реки Волги в её низовьях в районе г. Астрахани впервые были
проведены в 1909 г. М.Х. Элъдаровой, Сергеевой. Было зарегистрировано 400
таксонов водорослей.
Общая картина сезонной
динамики развития фитопланктона в Волге, уставленная в начале века довольно
близко совпадает с современной. По данным К.В.Горбунова (1976), в водоемах
дельты Волги, исключая подстепные ильмени было найдено 416 таксонов водорослей.
Пер"' те сведения фитопланктоне подстепных ильменей были получены
А.Ф.3иновьевым в 1937г. (Новожилова, 1987)
Отдельные сведения о
фитопланктоне подстепных ильменей дельты Волги опубликованы в небольшом числе
работ. В этих работах были даны лишь отрывочные сведения по интересующему
вопросу, выполненные к тому же на единичных водоемах. В настоящем разделе
сделаны попытки обобщить все имеющиеся многолетние данные по изучению
фитопланктона подстепных ильменей. (Антипчук, 1972).
В таблице 1 приведены
материалы по общей биомассе и доминирующей группе фитопланктона западных
подстепных ильменей, изученных в1983 г.
Таблица 1
Биомасса фитопланктона (В, г/м) разнотипных естественных ильменей западной части дельты Волги
| Название ильменя |
Площадь, га |
Средняя
глубина, м |
Биомасса |
Доминирующая
систематическая группа |
| Шушой |
300 |
1,2 |
24,27 |
синезел.,
диатомовые |
| Власов |
120 |
1.0 |
12.01 |
диатом.,
жчутковые |
| Большая Хошма |
200 |
0,9 |
3,22 |
диатом.,
зигнемовые |
| Табув |
120 |
0,8 |
2,53 |
диатом.,
зигнемовые |
| Уласты |
300 |
1,4 |
2,24 |
диатом.,
протококковые |
| Утусун |
180 |
1,0 |
13,68 |
диатом.,
протококковые |
| Соленый |
40 |
0,5 |
6,73 |
жгутиковые |
| Юнхара |
150 |
1,3 |
2,11 |
диатомовые |
| Бурата! |
180 |
1,5 |
3,96 |
диатом.,
синезеленые |
| Баркасный |
270 |
1,3 |
3,69 |
диатом.,
жгутиковые |
| Кривой |
150 |
1,0 |
2,71 |
диатом,
синезеленые |
| Долгий |
120 |
1,0 |
14,54 |
синезел.,
диатомовые |
| Яров |
100 |
0,6 |
5,90 |
волькосовые,
синезеленые |
| Большой Сарул |
250 |
1,1 |
11,28 |
диатом.,
синезеленые |
| Соленый - 2 |
60 |
0,3 |
0,17 |
улотриксовые,
синезел. |
| Кара - Булак |
250 |
0,9 |
17,87 |
синезеленые |
| Кальткун |
180 |
1,0 |
14,87 |
зигнемовые,
диатомовые |
| Задний Хагын |
200 |
1,3 |
1,98 |
диатомовые,
демидиевые |
| Булухта |
180 |
1,1 |
5,09 |
диатомовые,
жгутиковые |
| Большая Чада |
300 |
1,2 |
13,28 |
зигнемовые,
протококковые |
| Малая Чада |
60 |
0,9 |
23,54 |
диатомовые,
жгутиковые |
| Харнур |
100 |
1,0 |
8,34 |
диатомовые,
протококковые |
| Малый
Чапчалган |
100 |
1,0 |
10,54 |
синезеленые,
диатомовые |
Из данных таблицы 1
видно, что среднесезонные показатели общей биомассы фитопланктона в различных
ильменях не одинаковы. Из общего числа обследованных ильменей 38,5 %
низкопродуктивны (биомасса фитопланктона меньше 4 мг/л). Оставшиеся водоемы по
степени развития фитопланктона относятся к среднепродуктивным (биомасса 40-400
мг/л). Аналогичное соотношение в типах продуктивности ильменей получены на
основе съемки 1985 г. (табл.2).
Таблица 2
Концентрация
фитопланктона (В, г/м3) в разнотипных ильменях бассейна Северного
Каспия
| Район
исследования и название водоема |
Биомасса, г/м3
|
% от биомассы |
| зеленые |
диатомовые |
синезеленые |
эвгленовые |
пирофитовые |
| Казахстан |
|
|
|
|
|
|
| Куль – Тенес |
21,57 |
40,2 |
11,0 |
41,5 |
2,6 |
4,6 |
| Усек Куль |
185,8 |
21,0 |
70,8 |
4,6 |
3,6 |
- |
| Бидей Куль |
264,5 |
4,2 |
9,3 |
72,0 |
13,6 |
0,9 |
| Астраханская
область |
|
|
|
|
|
|
| Утусун |
189,1 |
0,36 |
86,9 |
9,9 |
2,9 |
0,1 |
| Глухой |
0,9 |
2,24 |
20,4 |
77,4 |
- |
- |
| Большая Чада |
2,5 |
15,8 |
38,9 |
40,5 |
4,9 |
0,9 |
| Шушой |
45,4 |
5,1 |
49,7 |
43,8 |
1,0 |
0,4 |
| Власов |
2,5 |
16,0 |
76,5 |
- |
4,0 |
3,5 |
| Большая Хошма |
11,4 |
2,1 |
75,4 |
17,4 |
3,4 |
1,7 |
| Уласты |
3,3 |
1,6 |
10,9 |
12,1 |
4,2 |
71,1 |
| Зубурунное |
1,5 |
21,4 |
58,3 |
2,7 |
- |
17,6 |
| Калмыкия |
|
|
|
|
|
|
| Олиничевское
водоохран. |
86,1 |
0,7 |
94,8 |
4,5 |
- |
- |
| Рыжковское
водоохран. |
9,7 |
7,2 |
0,5 |
76,6 |
8,7 |
5,9 |
| Красинский
банк |
3,3 |
5,7 |
86,9 |
7,1 |
- |
- |
Во всех обследованных
ильменях наблюдался разнохарактерный тип количественной динамики общей биомассы
фитопланктона. В большинстве случаев наблюдалось возрастание концентраций
водорослей от весны к лету. Так, в наиболее продуктивных ильменях рост биомассы
водорослей от весны к лету зафиксирован Чапчалгане.
Таблица 3
Концентрация водорослей в воде ильменя Большой Чапчалган
| Водоросли |
Сезоны года |
| Весна |
Лето |
Осень |
|
Мг/м3
|
% |
Мг/м3
|
% |
Мг/м3
|
% |
| Диатомовые |
206,9 |
17,1 |
2254,8 |
2,3 |
9356,5 |
4,6 |
| Протококковые |
220,8 |
18,2 |
574,9 |
0,5 |
475,9 |
2,4 |
| Зигнемовые |
0,49 |
0,04 |
- |
- |
- |
- |
| Улотриксовые |
75,4 |
6,2 |
- |
- |
13353,2 |
66,1 |
| Хризомонады |
7,41 |
0,6 |
- |
- |
- |
- |
| Вольвоксовые |
- |
- |
- |
- |
145,9 |
0,7 |
| Жгутиковые |
131,1 |
10,8 |
- |
- |
21,65 |
0,11 |
| Синезеленые |
568,8 |
46,9 |
97273,9 |
97,2 |
- |
26,1 |
| Всеговодорослей |
1210,9 |
100 |
100076,6 |
100 |
- |
100 |
Таблица
4
Концентрация водорослей в воде ильменя Малая Чада
| Водоросли |
Сезоны года |
| Весна |
Лето |
Осень |
|
Мг/м3
|
% |
Мг/м3
|
% |
Мг/м3
|
% |
| Диатомовые |
448,4 |
49,1 |
3651,8 |
5,6 |
3012,1 |
70,7 |
| Протококковые |
15,1 |
1,7 |
248,6 |
0,4 |
156,4 |
3,7 |
| Хризомонады |
2,9 |
0,3 |
958,2 |
1,5 |
61,1 |
1,4 |
| Вольвоксовые |
20,8 |
2,3 |
41,9 |
0,1 |
- |
- |
| Жгутиковые |
412,7 |
45,2 |
52053,3 |
79,5 |
992,4 |
23,3 |
| Синезеленые |
13,5 |
1,4 |
8502,7 |
12,6 |
37,1 |
0,9 |
| Всеговодорослей |
913,4 |
100 |
65456,5 |
100 |
4259,1 |
100 |
От высоких общих масс
фитопланктона весной - 57,9 г/м3, где господствующей формой была
синезеленая водоросль Oscillatoria planctonica
произошел спад биомассы к лету, который продолжался до осени. В летний период в
этом водоеме доминировали три вида Синезеленых: Merismopedia glauca, Anabaena sp., O. Limnetica и два вида зеленых: Sc. Quadricauda, P. Boryanum и два
вида диатомовых: N. gracilis, Surirella sp.
Осенью в составе диатомового
фитопланктона доминировали два вида: Pinnularia sp., Cocconeus placentua.
Таблица 5
|
Концентрация
водорослей в воде ильменя Культкун. |
| Водоросли |
Сезоны года |
|
| весна |
лето |
Осень |
|
|
Мг/м3
|
% |
Мг/м3
|
% |
Мг/м3
|
% |
|
| Диатомовые |
9353,7 |
22,1 |
353,4 |
24,8 |
681,9 |
90.2 |
|
| Протококковые |
222,6 |
0,5 |
215,9 |
15,2 |
40,7 |
5,4 |
|
| Зигнемовые |
30965,4 |
73,1 |
- |
- |
- |
- |
|
| Десмидиевые |
703,6 |
1,7 |
- |
- |
- |
- |
|
| Улотриксовые |
- |
- |
853,6 |
58,6 |
- |
- |
|
| Волъвоксовые |
968,9 |
2,3 |
- |
- |
- |
- |
|
| Жгутиковые |
37,9 |
0,09 |
- |
- |
33,3 |
4.4 |
|
| Синезеленые |
159,2 |
0,39 |
21,7 |
1,5 |
- |
- |
|
| Всего
водорослей |
42410,3 |
100 |
1426,6 |
100 |
755,8 |
100 |
|
Следует, однако,
отменить, что видовой состав доминирующих форм водорослей в этом водоеме
отличался от такового в ильмене Шушой. Весной доминировал один вид зигнемовых –Mongeotia sp., составлявший свыше 70 % биомассы всего фитоценоза. В
летний период доминировал один вид улотриксовых – Coeoleochaeta scutata, а осенью диатомовые: Denticula sp., Amphora ovalis.
(Сокольский, 1995)
Прямо противоположный тип
сезонной динамики общей биомассы фитопланктона наблюдался в ильмене Малая Лага.
(табл.6)
Таблица
6
| Концентрация
водорослей в воде ильменя Малая Лага. |
| Водоросли |
Сезоны года |
| весна |
лето |
осень |
|
Мг/м3
|
% |
Мг/м3
|
% |
Мг/м3
|
% |
| Диатомовые |
74422,2 |
12,2 |
38,8 |
0,08 |
7232,2 |
4,32 |
| Протококковые |
10362,1 |
17,1 |
6483,1 |
13,7 |
928,0 |
0,6 |
| Вольвоксовые |
3487,1 |
5,7 |
371,3 |
0,8 |
- |
- |
| Жгутиковые |
4035,7 |
6,6 |
7693,5 |
16,2 |
148,1 |
^.^^ |
| Синезеленые |
36629,9 |
58,5 |
32871,7 |
69,3 |
158919,9 |
95,1 |
| Всего
водорослей |
60937,9 |
100 |
47458,4 |
100 |
167227 |
109 |
В этом водоеме в отличие
от ильменей Шушой и Культкун при постоянно высокой биомассе фитопланктона к
осени наблюдался рост синезеленых. Господствующими формами были те же виды
синезеленых – Oscilatoria limosa, O. Limnetica и O. Pianctonica;
осенью их биомасса достигала 167,2 г/м3. Ильмень Горячинский при
более умеренных биомассах фатопланктона имел на протяжении сезона два пика - в
мае и августе. Весенне-летний пик был обусловлен развитием протококковых schroderie setigera и Sc. Opolionsis, а
осенний за счет синезеленых (О. limosa,
O. Pianctonica) и жгутиковых (Euglena sp., Lepocinelis sp.). (Сокольский,
1995)
Таким образом, типы
динамики фитопланктона во всем многообразии естественных ильменей различны и
могут быть представлены четырьмя формами: 1 - подъем биомассы от весны к лету с
последующим понижением к осени; 2-падение биомассы от весны к лету и далее к
осени; 3- непрерывный подъем биомассы с максимумом осенью; 4- двухвершинная
кривая (максимумы развития водорослей в весенне-летний и летне-осенний период).
Представляло интерес
рассмотреть влияние солености на биомассу, фитопланктона ильменей, (табл.7)
Таблица
7
Развитие фитопланктона в
ильменях в зависимости от их солености
| Водоем |
Биомасса 1г/м3
|
Соленость Мг/л |
Водоем |
Биомаса г/м3
|
Соленость МГ/Л |
| Кара- Булак |
17,87 |
64,91 |
Шушой |
24,3 |
50,4 |
| Кальткун |
14,87 |
74,24 |
Власов |
12,1 |
61.0 |
| Чапчалган |
|
350,0 |
Долгий |
14,54 |
159,5 |
| Кошата |
24,39 |
1415,0 |
Утусун |
13.68 |
3176,9 |
| Соленый |
0,91 |
27300,0 |
Соленый |
0,17 |
33200,0 |
Из приведенных в таблице
данных видно, что соленость воды в пределах 3 0/оо не влияет на
продуктивность фитопланктона. В пределах указанных выше концентраций солей
продуктивность фитопланктона ильменей, по всей видимости, определится другими
факторами, повышение солености воды ильменей до 30°/оо резко угнетает развитие
водорослей.
Известно, что
физиологическая активность водорослей зависит от их размеров (Лаврентьева,
1981). Размер организмов фитопланктона характеризует трофический статус
водоемов. (Лаврентьева. 1985).
Подавляющее большинство
форм водорослей в ильменях представлена крупными (свыше 10 мкм) особями. Такая
размерная структура доминирующих форм фитопланктона позволяет отнести
естественные ильмени к водоемам средней продуктивности. Следует подчеркнуть,
что для зоопланктона и белого толстолобика размерная структура фитопланктона ильменей
малопригодна для эффективного потребления. (Руденко,1995)
Колебания показателей
среднесезонной общей биомассы фитопланктона все они укладываются в пределы 1-40
г/м3, которые по классификации Г.Г.Винберга и В.П.Ляхновича,
характерны для низко- и среднепродуктивных водоемов.(Сокольский, 1995).
1.3.2
Состояние зоопланктона Западных подстепных ильменей
Первые работы,
посвященные фауне планктонных беспозвоночных Нижней Волги и её дельты,
появились еще в начале XX века (Боляхонцев,1901,1903).
В 40-х годах фауну
беспозвоночных в дельте Волги изучали В.С.Ивлев, М.И. Мухаревская,
А.Ф.Зиновьев. Наиболее обширные исследования в 1950 - 1970 гг. по зоопланктону
дельты и авандельты Волги выполнила А.А.Косова. Ею определено для этого района
218 таксонов коловраток, 86 ветвистоуеых и 53 веслоногих ракообразных. Для
основного русла Волги Е.К. Курашова приводит список, в котором отмечено лишь 57
таксонов, из них 26 коловраток, 14 ветвистоусых и 9 веслоногих ракообразных.
С целью более детального
изучения биологии зоопланктона в естественных ильменях в 1983-1987 гг.)
проводились исследования на данных водоемах разного типа (пресноводных,
солоновато водных, соленых). Материалы по общей численности зоопланктона,
изученных ильменей представлены в таблице 8.
Таблица
8
Общая численность
зоопланктона (тыс./м3) в разнотипных ильменях.
| Год |
Водоем |
Тип ильменя |
% от
численности |
| Численность |
Copepoda |
Cladocera |
Rotatoria |
| 1983 |
Власов |
Пресноводный |
156,18 |
83,9 |
6,2 |
9,9 |
|
|
Табун |
- |
181,4 |
66,3 |
26,7 |
0,1 |
| Улатсы |
Солоноватоводный |
138,6 |
68,8 |
13,6 |
14,9 |
| Соленый-3 |
Гиперсоленый |
0,3 |
100 |
- |
- |
| Кара-Булак |
Пресноводный |
395,8 |
30,7 |
46,6 |
24,7 |
| Кулькун |
- |
133,7 |
59,2 |
28,6 |
12,2 |
| Чапчалган |
- |
104,7 |
69,7 |
9,4 |
20,1 |
| Хошата |
Солоноватоводный |
109,6 |
46,7 |
6,1 |
47 |
| Соленый-4 |
Гиперсоленый |
0,4 |
100 |
- |
- |
| Долгий |
Пресноводный |
462,9 |
39,9 |
32,9 |
26,8 |
| Задний Хагын |
Солоноватоводный |
51,6 |
72,1 |
19,1 |
8,8 |
| Большая Хошма |
- |
93,1 |
67,2 |
30,5 |
2,4 |
| 1984 |
Кара-Булак |
Пресноводный |
102,3 |
37,7 |
19,9 |
37,3 |
| Кулькун |
- |
124,1 |
59,7 |
7,7 |
25,3 |
| Чапчалган |
- |
113,6 |
62,7 |
14,3 |
20,2 |
| Хошата |
Солоноватоводный |
73,4 |
50 |
12,6 |
37,4 |
| 1985 |
Кара-Булак |
Пресноводный |
225,1 |
62,7 |
45,6 |
37 |
| Галга |
- |
158,9 |
30,7 |
5,5 |
31,8 |
| Хошата |
Солоноватоводный |
558,9 |
49,7 |
4 |
65,4 |
| 1986 |
Галга |
Пресноводный |
221 |
56,7 |
6 |
44,3 |
| 1987 |
Галга |
- |
92,5 |
|
10,2 |
33,1 |
Из данных таблицы видно,
что численность зоопланктона в естественных ильменях не превышает 600 тыс./м3,
что характеризует данные водоемы, как мезоолиготрофные. (Сокольская, 1995).
Следует отметить
тенденцию к уменьшению общей численности зоопланктона в солоноватоводных и
особенно гиперсоленых ильменях, по сравнению с таковой пресноводных водоемов. В
большинстве случаев в независимости от типа водоема (пресноводный или
солоноватый) в составе зоопланктона преимущественно развивались веслоногие раки
и коловратки. По сравнению с этими двумя группами, развитие ветвистоусых
ракообразных было выражено в меньшей степени. (Сокольский,1973)
По числу видов в ильменях
также доминировали коловратки, второй по значению группой были ветвистоусые
ракообразные и веслоногие раки. Из коловраток наиболее часто встречались
следующие формы: Asplanchna priondata, A. brightwelli, Brachyonus caliciflorus amphiceros, B. diversicornis, B. angularis, B. quadridentatus melhini, Euchlanis dilatata, Filinia longiseta, K. guadrata
quadrata.
Довольно часто им
сопутствовали: Brachyonus caliciflorus anuraeciformes, Euchlanis ircisa, Keratella coclilearis, Polyarthra sp., Synchaeta sp., Lecane luna, Platyas patulus
patulus, B. urcens, B. q. clunioorbicularia,
B. q. ancylognathus. К
формам встречавшимся эпизодически, относятся: Brachyonys caliciflorus anuraeciformes, Lecane bulla, Philodinidae sp., Festudinella patina.
Среди ветвистоусых
ракообразных в зоопланктоне наиболее часто встречались: Alona rectangular, Alonella nana, B. longirostris, Pleuroxus sp., Moina macroscopa, Scapholeberis mucronata.
В весенний период наряду
с ними в планктоне появлялись: Hiocryptus acutifrons, Sida crystalline, pleuroxus sp. В летне-осенний период эти организмы выпадали из зооценоза,
а на смену им приходили другие формы: Graptolberis mucronata, Daphnia longispina, Leydigia acantocercoides.
Веслоногие раки в
пресноводных ильменях были представлены науплиальными и копеподитными стадиями
циклопоида и каланоида. Среди определенных форм Calanoida наиболее часто встречались Eurytemora affinis и E. velox.
1.3.3.
Биомасса зоопланктона естественных ильменей
Основной задачей этого
раздела является уточнение трофического статуса естественных ильменей на основе
изучения биомассы зоопланктона. В таблице приведены данные по общей биомассе
зоопланктона и составляющих его групп в различных ильменях. (Сокольский, 1992)
Таблица 9
Общая биомасса
зоопланктона (г/м3) в разнотипных естественных ильменях
| Год |
Водоем |
Тип ильменя |
% от
численности |
| Численность |
Copepoda |
Cladocera |
Rotatoria |
| 1983 |
Власов |
Пресноводный |
0,57 |
77,1 |
17,5 |
3,5 |
|
|
Табун |
- |
0,96 |
57,8 |
40,9 |
1,1 |
| Улатсы |
Солоноватоводный |
0,71 |
80,3 |
15,5 |
4,2 |
| Кара-Булак |
Пресноводный |
0,07 |
87,14 |
5,7 |
10 |
| Кулькун |
- |
1,39 |
70,6 |
30 |
0,3 |
| Чапчалган |
- |
0,55 |
90,9 |
3,6 |
5,5 |
| Хошата |
Солоноватоводный |
0,26 |
76,9 |
3,8 |
19,2 |
| Долгий |
Пресноводный |
3 |
42,7 |
45,8 |
11,5 |
| Задний Хагын |
Солоноватоводный |
00,39 |
70,9 |
27,8 |
1,3 |
| Большая Хошма |
- |
0,37 |
56,1 |
43,1 |
0,8 |
| 1984 |
Кара-Булак |
Пресноводный |
0,39 |
57,3 |
1,5 |
41,2 |
| Кулькун |
- |
0,88 |
90,8 |
2,3 |
6,6 |
| Чапчалган |
- |
0,67 |
70,9 |
0,7 |
28,3 |
| 1985 |
Кара-Булак |
Пресноводный |
0,98 |
65,5 |
21,9 |
12,5 |
| Галга |
- |
0,87 |
74,5 |
24,9 |
14,2 |
| Хошата |
Солоноватоводный |
2,58 |
89,9 |
3,1 |
7 |
| 1986 |
Галга |
Пресноводный |
0,73 |
65,5 |
3,2 |
31,2 |
| 1987 |
Галга |
- |
0,73 |
58,2 |
6,4 |
35,5 |
Из данных таблицы 9
видно, что в подавляющем большинстве естественных ильменей общая биомасса
зоопланктона не превышала 0,7 – 0,9 г/м3, что соответствует таковой
олиго-, мезотрофных озер. Среди трех основных групп зоопланктона в подавляющем
большинстве ильменей доминировали веслоногие ракообразные. (Сокольский, 1995).
В сезонной динамике общей
биомассы зоопланктона следует отметить, что наибольший пик её наблюдался весной (мае), затем она
уменьшалась (июнь) и вновь возрастала в конце периода наблюдений (августе).
Такой же характер сезонной динамики общей массы зоопланктона наблюдался в
ильмене Галга в 1987 (Табл.10).
Таблица 10
Сезонная динамика общей
биомассы и массовых видов зоопланктона ильменя Галга (1987 г.)
| Вид
зоопланктона |
Биомасса,
мг/м3 |
| 20.05. |
18.05. |
1.07. |
16.07. |
5.08. |
| Cyclopoida nauplii |
180,0 |
124,8 |
16,0 |
151,8 |
205,2 |
| Cyclopoida copepoditi |
44,0 |
28,1 |
3,5 |
22,2 |
16,0 |
| Cyclopoida sp. |
125,9 |
731,2 |
4,0 |
12,1 |
35,6 |
| Asplanchna brightwelli |
288,0 |
- |
20,8 |
- |
- |
| Asplanchna priodonta |
1216,0 |
- |
- |
- |
- |
| Asplanchna herricki |
- |
- |
- |
32,0 |
- |
| Brachionus calamphiceros |
52,0 |
- |
- |
85,8 |
46,8 |
| Moina macrocopa |
- |
- |
11,2 |
- |
- |
| Общая
биомасса: |
1595,1 |
952,9 |
66,1 |
329,8 |
326,9 |
Из данных таблицы видно,
что пик общей биомассы зоопланктона наблюдался в июле. Общим было то, что от
мая к июню общая биомасса зоопланктона уменьшалась. Сравнивая данные табл.10 и
табл.11 видно, что во всех изученных ильменях в зооценозе доминировали
представители веслоногих ракообразных и коловраток. (Сокольский,1995)
Таблица 11
Динамика обшей биомассы и
массовых видов зоопланктона ильменя Галга (1986 г.)
| Вид
зоопланктона |
Биомасса,
мг/м3 |
| 14.05. |
17.06. |
3.07. |
16.07. |
7.08. |
| Cyclopoida nauplii |
200,0 |
221,2 |
519,6 |
477,6 |
71,5 |
| Cyclopoida copepoditi |
57,0 |
50,8 |
75,0 |
111,0 |
40,8 |
| Cyclopoida sp. |
301,2 |
15,2 |
109,1 |
- |
- |
| Brachionus calanphiceros |
65,6 |
- |
- |
- |
- |
| Euchlanis dilatata |
88,0 |
- |
- |
- |
- |
| Asplanchna priodonta |
- |
176,0 |
- |
108,0 |
- |
| Asplanchna brightwelli |
- |
- |
- |
108,0 |
- |
| Synchaeta stylata |
- |
- |
139,9 |
- |
- |
| Brachionus amphiceros, amph. |
- |
- |
- |
- |
31,7 |
| Общая
биомасса: |
856,6 |
538,6 |
1032,8 |
1051,8 |
172,9 |
В заключение отметим, что
по численности и биомассе зоопланктона естественные ильмени дельты Волги можно
отнести к олиго-, мезотрофным водоемам. (Сокольская, 1981)
В составе зоопланктона
ведущую роль играют мелкие формы: науплии и копеподиты веслоногих ракообразных,
а также коловратки. На основе изложенного можно предположить, что рыбы с
фильтрационным типом питаний (белый и пестрый толстолобики, а также, веслонос)
могут получать здесь преимущество перед рыбами с другим типом питания (карпом).
(Сокольский, 1995).
1.4.
Искусственное повышение биопродуктивности ильменей
Органические и
минеральные удобрения, вносимые в ильмени повышают естественную рыбопродуктивностъ
вследствие последовательного развития отдельных звеньев пищеводу цепи:
бактерий, фитопланктона, зоопланктона, бентоса. Удобрение водоемов приводит к
обогащению воды биогенными элементами, что способствует максимальному развитию
первичной продукции. Интенсивное развитие бактерий и фитопланктона, как
первичного звена трофической цепи, сопровождается увеличением численности
зоопланктона и бентоса, для которых первые служат пищей. Развитие фитопланктона
приводит к улучшению кислородного режима прудов. Кроме того, фитопланктон
является пищей для рыб. (Рыжков, 1973; Баранов, 1972, 1974; Баранов, Салазкина,
1969).
Минеральные удобрения
содержат биогенные элементы, дозировка и влияние которых на повышение
продуктивности водоемов определяется достаточно точно. Органические удобрения
значительно сложнее и разнообразнее. Их состав зависит от происхождения
(Бурдиян, Мухачев, 1975).
Эффект использования
удобрений определяется температурой вод активной реакцией, газовым составом,
проточностью, составом почвы, а также видовым составом выращиваемых рыб
(Егоров, 1969; Стеффенс, 1985).
Органические вещества,
находящиеся в воде во взвешенном состоянии, перифитон, бактериопланктон,
которые находятся на детрите и на сестоне, при их соединении становятся
дополнительной пищей рыб. Роль естественной пищи в питании прудовых рыб очень
велика. Естественная пища способствует обогащению неполноценного комбикорма
различными питательными веществами (Методическое пособие., 1974).
В прудах постоянно
происходит процесс размножения, развития и отмирания гидробионтов. В процессе
жизнедеятельности гидробионтов, в том числе и рыб, происходит накопление
остатков несведённого корма, остатков метаболизма. Много различных веществ
органического и неорганического происхождения попадает в пруд извне, а также
оседает на дно: в результате накапливаются донные осадки, ил, иловые отложения.
За год в пруду накапливается слой толщиной 0,5 - 2 мм. Иловые отложения играют
существенную роль в жизни пруда. В них происходит развитие бентических
организмов, для которых ил служит пищей, идет интенсивный процесс разложения
(минерализации) органических веществ, в котором основную роль играют бактерии.
Процесс минерализации протекает при значительном потреблении кислорода,
растворенного в воде. Поэтому в интенсивно эксплуатируемых прудах, в которые в
большом количестве вносят удобрения, часто возникают заморные явления.
(Мухачев, 1971, Решетников, Попова, Стерлигова, 1982).
Бактерии в огромном количестве
развиваются не только в илах, но и в толще воды, и на сестоне. Разлагая мертвое
органическое вещество, они очищают водоем. Бактерии являются пищей
зоопланктона. В то же время слишком интенсивное развитие бактериопланктона
нежелательно, поэтому в пруды периодически вносят хлорную или негашеную известь
для частичного уничтожения бактерий (Методические указания.., 1990).
В поисках донных
организмов (бентоса) рыбы - бентофаги взмучивают ил, что происходит, в
основном, в светлое время суток. Ночью, взмученный ил вновь оседает. Такое
перемешивание дает бактериям получать в необходимом количестве растворенный в
воде кислород, что приводит к ускорению минерализации органических веществ и
очищению водоема. (Рыжков, 1983,1987).
Чтобы увеличить
количество естественной пищи в водоемах, разработаны методы воздействия на
водную экосистему. Одним из методов, создающих благоприятную среду для развития
бактерий, является внесение в водоемы органических удобрений. Бактерии являются
пищей для беспозвоночных ракообразных. Другим методом является внесение азотных
и фосфорных удобрений, являющихся питательными веществами для планктонных
водорослей, которые в свою очередь являются пищей для зоопланктонных
ракообразных и растительноядных рыб. Большой эффект по увеличению естественной
кормовой базы и рыбопродуктивности дает метод интродукции (вселения) в
выростные пруды Daphnia
magna - наиболее крупного и
высокопродуктивного представителя низших ракообразных (зоопланктона). Чистую
культуру Daphnia magna вносят в пруды при залитии. Вместе с интродуцируемыми
животными вносят и корм для них - кормовые дрожжи и органические удобрения. В
пруду Daphnia magna быстро развивается и заселяет водную толщу, подавляя
развитие других, менее продуктивных животных, что позволяет увеличить естественную
рыбопродуктивность выростных прудов на 100 - 400 кг/га и более (Руденко,
Гвоздев, 1977; Рыжков,1973; Стерлигов, 1981).
1.6.
Пути
управления биоресурсами ильменей
Концепция эффективного
использования биологической противности ильменей разработана на основе
детальных знаний экологии изучаемого объекта. (Сокольский,1992).
Расчеты промыслового
запаса рыб, выполненные по кормовой базе, показали, что рыбопродуктивность
ильменей в среднем составляет 350 кг/га. Определения, проведенные на ильменях,
связанных непосредственно с р. Волгой, дают более высокую величину – 1000
кг/га. Расчет промыслового запаса на основе двух способов показывает, что с 100
тыс. га можно получить 35-100 тыс. т рыбы. В настоящее время объем добычи
составляет 300 т, что соответствует 0,8— 0,3 % интенсивности вылова.
(Сокольский, 1992).
Следовательно, увеличение
объемов добычи рыбы на ильменях - одно из важнейших направлений их
рационального использования. Помимо рыбы, в западных и восточных подстепных
ильменях достаточно большие запасы раков. В последние годы добыча этого объекта
неуклонно растет. Так, если в 1985 г. из ильменей было выловлено 5 т раков, то
в 1990г.- уже 20,9 т. Максимальный улов составлял 22,5 т (1989 г.).
Ракопродуктивные ильмени Монетный, Пальминный, Бугас, Туркменка, Байковский,
Карбута, а также практически все водоемы Красноярского района, требуют особой
охраны и организации на них рационального промысла. Контроль за промыслом раков
со стороны органов рыбоохраны важен потому, что как показывают наблюдения, бесконтрольный
ежегодный отлов раков из одних и тех же ильменей приводит к потере их
ракопродуктивности. Так, в ильмене Бугас в 1985 г. было добыто 4 т раков, в
1988 и 1989гг.- 1,5 т., в 1990г. только 0,3 т. (Сокольскйи,1992).
В последнее время
промышленность обратила внимание на возможность добычи из ильменей сапропеля.
Как показали исследования, наиболее качественное сырье в проточных ильменях, в
замкнутых водоёмах качественный состав сапропеля значительно хуже. Предварительное
обследование показало, что только из 3-х ильменей Восточненского водного тракта
(Лата, Сарул, Алчинский) можно
добыть около 20 млн. т сапропеля. В ценах 1992 г. рентабельность составит 315%,
чистая ежегодная прибыль – 7,9 млн. руб. Учитывая, что в настоящее время
существует 7 водных трактов с общей площадью водного зеркала 40 тыс.га из этих
водоемов можно добыть свыше 100 млн.т сапропеля, что позволяет получить около 1
млрд.руб. чистой прибыли. (Сокольский, 1992).
Повышение биологической
продуктивности ильменей возможно только при решении такой важнейшей
стратегической задачи, как эффективная мелиорация. В настоящее время площадь
"Каспия "увеличилась на величину, равную Азовскому морю. В результате
усилившегося подпора вода со стороны моря существующие каналы-рыбоходы начали резко
заиливаться. Доля плоскостного стока неуклонно растет и составляет в настоящее
время до 70 % общего объема. Заработали старые жилки, и рыба начала скатываться
в основном банчинами. Таким образом, налицо эффект, который условно назван
"инфаркт дельты". Поступление воды в ильмени ограничивается еще и
огромным числом завалок, до сих пор, не ликвидируемых колхозами и совхозами.
Следовательно, уже сейчас необходимо приступить к разработке Генеральной схемы
мелиорации дельты р. Волги, учитывающей изменившуюся экологию Каспийского моря.
(Сокольский, 1992).
Не потеряла актуальности
и проблема превращения части потерявших рыбохозяйственное значение ильменей в
озерные товарные рыбоводные хозяйства. Биотехнология выращивания карповых и
осетровых рыб на этих водоемах разработана достаточно полно. Следует заметить,
что организация таких хозяйств на площади, свыше 100 тыс. га, может дать
промышленности до 21 тыс.т карповых и около 3 тыс.т. осетровых (включая
веслоноса). (Федорченко, Новоженин,3айцев,1992).
1.7.
Состояние запасов рыб в западно-подстепных ильменях и перспективы их промысла
Заход рыб в
западно-подстепные ильмени связан с водностью в период весеннего паводка. Так,
в маловодном 1989г. выловлено 37,66т., в маловодном 1995 г. - 334,95 т.
Среднегодовые промысловые уловы рыб в последнее десятилетие составили 251,40 т.
Доля крупного частика в уловах колебалась от 26,6 % в маловодном 1996 г. до
73,7% - в многоводном 1990 г. В уловах крупного частика большую долю составил
сазан - в среднем 13,7%, меньшее значение имели лещ - 10,5, щука - 7,9, сом -
2,7, судак -1,2%. Вылов рыб этих видов положительно коррелирует с объемом
весеннего паводка. (Сокольский, 1997). Рыбы - акклиматизанты присутствуют в
уловах не ежегодно. Среднегодовой вылов белого амура составляет 0,35 т, пестрого
и белого толстолобиков -12,41 т. Неустойчивость уловов растительноядных рыб
связана с нестабильностью их воспроизводства.
Среди мелких частиковых
рыб преобладают караси - 14,2%. При этом доля в общем улове серебряного карася
с годами растет, а золотого - снижается; основными видами являются сопа -10,2%;
красноперка - 9,9 и густера - 9,0%; доля окуня снизилось да 7,2%;
незначительными стали среднегодовые уловы воблы - 3,5, линя - 2,1, язя - 0,2%
общего улова (в последнее время язь вообще не встречается).
Наблюдается рост уловов
серебряного карася и сопы. Эти виды в ближайшие годы будут доминирующими. Вылов
промысловых рыб будет зависеть от водности во время весенних паводков: при
обильном половодье он может достигать 0,35 тыс.т, маловодном - 0,05 тыс.т. (Сокольский,
1997).
В последние годы
маловодных паводков ожидается перераспределение структуры уловов в пользу
малоценных рыб, в первую очередь, серебряного карася и сопы. Вместе с окунем,
густерой и красноперкой эти виды могут составлять 80 % уловов.
Повышению рыбохозяйственной
значимости западно-подстепных ильменей будут способствовать рациональный режим
весеннего паводка и своевременная "реконструкция" ихтиофауны, в
частности, вселение веслоноса, пиленгаса и растительноядных рыб. (Сокольский,
1997).
Глава 2.
Эколого-географическая характеристика западно-подстепных ильменей
2.1.
Происхождение Ильменей
Из общей площади дельты
Волги, 24292 квадратных километра, Западные подстепные ильмени занимают 5907 м2
Е.Ф. Белевич (1963)
выделяет этот район как самостоятельную часть дельтовой области реки Волги.
Рассматриваемый район отличается своеобразными природными условиями - климатом,
водным режимом, почвенным покровом и т.д. (Сокольский, 1995). Район западных
подстепных ильменей, как и всего Каспийского моря и Дельты Волги, представляет
собой часть Прикаспийской бесточной котловины, образование которой относят к
третичному периоду. Уровень Каспия подвергался значительным многократным колебаниям
в результате трансгрессий. Самой большой в четвертичном периоде была хвалынская
трансгрессия (около 10 тысяч лет тому назад), при которой отложились толщи
шоколадных хвалынских глин. (Ушаков, Шукшина, Тимофеева, 1995).
В низких межбугровых
котловинах отмечается накопление илистых осадков в результате ежегодных
половодий, приносящих илистые частицы. В таких понижениях между буграми Бэра и
сформировались подстепные ильмени. Важной особенностью всех бугров Бэра (они были
описаны академиком К.М.Бэром в 1856 г.) является их строгая ориентировка с
востока на запад. Соответственно ориентированы и котловины ильменей.
(Сокольский, 1995).
Средняя высота бугров Бэра
10-12 м, ширина 200- 500 м, а длина варьирует от 200 м до 10-20 км. Расстояние
между буграми от 200м до 1 км, иногда и больше. Между буграми встречаются как
занятые водой, так и высохшие ильменные западины. И бугры Бэра, и,
соответственно, ильмени, уходят от Волги далеко на запад (более 100 км от
основного русла Волги) и там примыкают к полупустынной степи Калмыкии.
(Сокольский,1995).
2.2.
Типология западных подстепных ильменей
Южнее Волгограда от Волги
отходит рукав - река Ахтуба, которая течет около 500 км почти параллельно
Волге, то, удаляясь от нее до 30 км, то, приближаясь к ней до 7 км. Волга и
Ахтуба образуют обширную пойму со множеством озер, протоков, ериков и рукавов.
Скорость течения в Волге в меженное время невелика (0,3 - 0,5 м/с). Ахтуба
многоводна только в период весеннего половодья, летом на перекатах сильно
мелеет. Южной границей поймы считают р. Бузан. В этом месте пойма резко
расширяется и переходит в дельту. Дельта подразделяется на надводную и
подводную. Площадь первой - 18949 км2, второй - 5343 км2.
Основные исследования
осуществлялись в зоне Западных подстепных ильменей, являющихся составной частью
дельты Волги. Из общей площади дельты 24292 км2 Западные подстепные
ильмени занимают более 5,9 тыс.км2. Е.Ф. Белевич (1963) выделяет
этот район как самостоятельную часть дельтовой области реки Волги.
Несмотря на близость
Каспийского моря и обильное волжское половодье, весь этот район в климатическом
отношении близок к полупустыням. Климат здесь резко континентальный. Обычно
имеет значительный недостаток атмосферных осадков. В среднем, за год их
выпадает от 133,2 до 233,9мм. Началом зимы здесь считается примерно середина
декабря, когда наступают устойчивые морозы. В конце февраля начале марта
начинается весна, а во второй половине апреля кончаются заморозки и начинается
лето. Лето продолжительное, жаркое и сухое. Средняя температура воздуха в июле
- от 24 до 27.°С, максимальная - до +40°С. Температура воды в июле-августе в
ильменях держится на уровне +25-27°С, достигая в отдельные дни 30°С. До конца
сентября температура воды в озерах не опускается ниже +20°С.
Испарение в районе
ильменей превышает количество выпадающих осадков почти в 10 раз, и поэтому
многие ильмени, расположенные на границе с Калмыкией, к осени пересыхают.
(Чуйкой, 1984). Некоторые исследователи разделяют район Западных подстепных
ильменей на, две части: западную, или степную, и восточную, или прирусловую
(Брюшков, 1951). Неоднократно делались попытки классифицировать Западные
подстепные ильмени. Так, Н.Л.Чугунов (1923) все ильмени делит по степени
проточности на две группы. В.И. Мейснер (цитируется по Плюснину, 1958),
классифицирует ильмени по генетическому признаку и выделяет три группы. (Сокольский,
1995).Позднее была предложена классификация ильменей, по которой все они были
разбиты на 3 большие группы:
1. Постоянные, то есть
никогда не пересыхающие, глубокие ильмени (площадь таких водоемов в настоящее
время более 140 тыс. гектаров);
2. Постоянные
непересыхающие мелководные (общая площадь 50-75 тыс. га);
3. Пересыхающие в
меженное время (площадь 200-300 тыс. га – западная оконечность ильменей).
Данная классификация, не
будучи детализированной, позволяла описать большие группы водоемов, существующие
в настоящий период, на основе главных показателей их гидролого-гидрохимического
режима. (Сокольский, 1995).
Впоследствии появилась
детальная классификация водоемов Волго-Каспийского района предложенная Ю.С.
Чуйковым и др.(1994), в которой приводится и типология ильменей. По
предложенной авторами классификации все разнообразие водоемов зоны Западных
подстепных ильменей объеденено в 9 больших типов. С учетом сделанных дополнений типология Западных подстепных
ильменей в методологическом понимании указанных выше авторов представлена в
таблице 13.
Рассматривая типологию
водоемов, представленную выше, следует отметить, что не все из представленных
типов ильменей имеют рыбохозяйственное значение. Наибольшую ценность
представляют безусловно, мелиорированные или искусственные водоемы, которые
дают свыше 90 % вылавливаемой в этом районе рыбы. Практически не имеют
рыбохозяйственного значения все другие представленные в классификации типы
озер, за исключением пресноводных глубоководных ильменей водохранилищ и водотоков,
входящих в состав водных трактов. (Сокольский, 1995).
Таблица
13
Типология Западных подстепных
ильменей дельты Волги
| Зональные
территориальные единицы |
Основные типы
водоемов |
Типы водоемов |
| Постоянные или
периодические пересыхающие озера в межбугровых понижениях и питающие их
протоки |
Пресные или
солоноватые ильмени |
1. пресные или
слабосоленые глубоководные ильмени с прибрежными тростниково-рогозовыми
зарослями (изучено 18 водоемов) |
| 2. пресные или
слабосоленые ильмени с плавневыми тростниково-рогозовыми зарослями (изучено
20 водоемов) |
| 3. соленые
мелководные ильмени (изучено 4 водоема) |
| Ультрагалинные
ильмени |
4. мелководные
ильмени с отложением осадочной соли (изучено 4 водоема) |
| Русловые
водоемы |
5. дельтовые
протоки, питающие системы ильменей (изучено 10 протоков) |
| 6. протоки,
соединяющие системы ильменей (изучено 10 протоков) |
| Мелиорированные
или искусственные водоемы |
7. Пресные
водоемы, мелиорированные для рыборазведения: выростные и нагульные карповые
пруды, нерестово-выростные хозяйства, осетровые выростные и нагульные пруды,
озерные товарные рыбоводные хозяйства (изучено 120 водоемов) |
| 8. Временные водоемы (рисовые чеки, оросительные каналы, сбросные
водоемы) (изучено 7 водоемов) |
|
|
9. ильмени – водохранилища и водотоки, ильмени – отстойники (изучено 7
водоемов) |
2.3.
Классификация ильменей
Неоднократно делались
попытки классифицировать подстепные ильмени. И.И. Плюснин (1958) дает более подробную
классификацию. Он выделяет следующие группы ильменей: 1) Высохшие ильмени –
мокрые солончаки; 2) Высыхающие ильмени с горько-соленой водой, удаленные от
моря и Волги и незатопляемые во время половодья; 3) Ильмени, затопляемые не
ежегодно, периодически, при максимальном подъеме уровня полых или вагонных вод,
с солоноватой водой; 4) Цепи озер, разделяющих между собой незначительными
перемычками, с затрудненным водообменом, с повышенной концентрацией солей в
воде удаленных пунктов; 5) Ильмени удлиненные, в виде заросших ериков, с
затрудненным водообменом, с повышенной концентрацией солей в воде удаленных
пунктов; 6) Широкие ильмени с пресной водой, беспрепятственно затопляемые при
всяком подъеме уровня воды, непосредственно примыкают к опресненной зоне моря
или рукавов реки Волги; 7) "Култуки" с пресной водой - начальная
стадия ильменей, ряд морских заливов, отражающие все изменения уровня
опресненной зоны моря. (Сокольский, 1992)
В.И. Мейснер классифицирует
ильмени по генетическому признаку: 1. Лагунные, 2. Култучные или морские. 3.
Ильмени речного происхождения.
Из них именно лагунные
ильмени располагаются в межбугровых понижениях, имеют вытянутую в широтном
направления форму, которая определяются ограничивающими их буграми. Среди них
имеются как проточные ильмени, соединенные с рекой, так и обособленные, причем
последние составляют подавляющее большинство. (Сокольский, 1992)
Среди последних имеются
ильмени, подвергающиеся влиянию вод весеннего разлива, и ильмени, у которых
приток свежей воды отсутствует даже в период половодья. Они легко заселяются.
Ильмени речного происхождения образуются из
отмирающих рукавов и стариц. Эти ильмени в своем большинстве постоянны, причем,
проточность сохраняется долгое время.
По морфометрическим
характеристикам подавляющее большинство Западных подстепных ильменей однородно.
Единичные водоемы имеют площади от 100 до 600 гектаров и среднюю глубину около
одного метра.
Интенсивная хозяйственная
деятельность человека в этой зоне изменила генезис этих водоемов. Часть
ильменей заполняется водой принудительно через систему водных трактов.
Вблизи от реки Бахтемир и
её протоков ильмени густо зарастают водной и водно-береговой растительностью,
вода в них не соленная. В прибрежье этих ильменей развиты заросли тростника (phragmites communes) или рогоза (Typha angustifolia).
Чем дальше к западу от р.
Бахтемир, по мере приближения к степным пространствам, вода в ильменях
становится солоноватой на вкус или совсем соленой. Развитие растительности
ограничивается. Вместо зарослей тростника и рогоза, берега опоясываются
бордюром различного рода солянок. В результате очень сильного испарения и
незначительного пополнения свежей водой, воды в таких ильменях становится
меньше, концентрация солей сильно возрастает и ильмени превращаются в соленые
озера или, пересыхая, в солончаки.
Для выяснения влияния
солености на гидрохимический режим ильменей, они исследованы по1-линии (Власов,
Табун, Уласты, Соленый-3); 2-я линия (Кара-Булак, Кулъткун, Чапчалган, Хошата,
Соленый -4), в которых они расположены последовательно по мере возрастания
общей минерализации воды. Концентрация гидрохимических компонентов, как в
ильменях одной линии, так и между линиями, в весенний период различалась
незначительно, за исключением солёности, которая закономерно увеличивалась.
(Сокольский, 1992).
Замкнутые ильмени с
грунтовым, питанием (Уласты) характеризуются возрастанием солености от весны к
лету, снижением концентрации фосфора, тогда как по биогенным элементам и
органическому веществу наблюдается картина, аналогичная слабопроточочным
ильменям. (Галасун, 1971).
Необходимо отметить, что
количественные характеристики процесса изменения гидрохимического состава воды,
происходящие от весны к лету, в ильменях, расположенных по линиям, резко
отличаются. Наиболее интенсивно процессы перестройки гидрохимического режима
проходят в ильменях Власов, Кара-Булок, расположенных вблизи водоподающих
рукавов. (Сокольский, 1992).
Для них характерно
большое расселение (до величин 70-80 мг/л), значительное увеличение
концентраций валового фосфора в 2,5 раза (Р. органического - в 3,5 раза), кремния
в 6 раз, возрастание окисляемости в 3 раза На других ильменях (менее
обводненных) аналогичные процессы проходят менее интенсивно. (Сокольский,
1992).
Для весенней воды
ильменей отношение азота к фосфору близко к 6, для летней - 4.1, осенней - 2.4,
для речной воды, питающей ильмени, это отношение постоянно и близко к 10,8. Эти
цифры свидетельствуют о двух важных моментах:
1.Речные воды не являются
определяющим фактором формирования биогенного режима ильменей;
2.Весной отношение азота
к фосфору, близкое к 6.0, соответствует стехиометрическому в планктоне. Поэтому
генетически азот и фосфор могут быть квалифицированы как регенерировавшие из
органического вещества в процессе минерализации его в осенне-зимний период;
Таким образом, следует
вывод о замкнутом круговороте (грунт-вода -грунт), как решающем факторе
формирования гидрохимического режима западных подстепных ильменей. Характер
обводнения ильменей в весенне-летний период оказывает воздействие на их
гидрохимический режим, в основном, вследствие механического перемешивания ила и
выброса его в воду, а также расселения. Роль пресных вод как поставщика
биогенных элементов незначительна. (Сокольский,! 992).
2.4.
Фнзико-географическсое положение Западных подстепных ильменей
Западные подстепные
ильмени расположены к западу от основной, центральной системы рукавов Волги. За
восточную границу распространения ильменей принимают обычно р. Бахтемир, но
правильнее было бы считать р. Хурдун, так как по естественным особенностям этот
участок (между Бахтемиром и Хурдуном) стоит ближе к центральной дельте. (Белевич,1958).
Вся область западных подстепных
ильменей в климатическом отношении представляет полупустыню с весьма малым
количеством атмосферных осадков (не свыше 180 мм в год), выпадающих летом в
виде кратковременных ливней. Средние летние температуры велики (до 25,3°). Зима
холодная, но короткая; огромная испаряемость (до 700 мм в год) и незначительный
снежный покров. Последний не всегда и не везде покрывает эту область и держится
не более двух месяцев в году. (Белевич,1958).
Область прорезана
множеством ильменей, связанных на севере с притоками Волги, на юге Каспийским
морем. Ильмени шире на востоке. Здесь они связаны с питающими их водоемами
(морем и рекой) в течение всего года. (Белевич.1958).
Наиболее значительная
часть ильменей сообщается с рекой только во время половодья или морян. Наконец,
есть ильмени, которые уже потеряли, связь с рекой и в настоящее время или
обсыхают или осолоняются. Отделение ильменей от питающих их водоемов особенно
интенсивно происходит в последнее время в связи с падением уровня Каспийского
моря. Дно ильменей плоское, слабо повышающееся в направлении берегов. Только в
центральной части водоема наблюдаются как бы слабые борозды — тальвеги, по
которым и осуществляется обмен вод между соседними ильменями, еще связанными
узкими протоками. Глубина ильменей
не превышает 2,0 м. Ильмени на востоке достигают 2 км и более ширины и в
направлении на запад суживаются до ширины протока – ерика а затем исчезают
совершенно. (Белевич, 1958).
В центральной части
описываемой области ильмени занимают до 20% всей поверхности, в паводок -
40%.(Белевич,1958).
Весь район западных
подстепных ильменей распадается по рельефу, на две отличные друг от друга части
— западную, или степную, и восточную, или ильменную, граница между которыми
условно проходит по меридиану 47° 15' в. д.. В направлении с востока на запад
количество понижений, занятых водой, уменьшается, поверхность степи делается
слабо увалистой, приобретая вид волнистого плато. Расчлененность рельефа
восточной части территории ильменей объясняется постоянным взаимодействием с
питающими их внешними водоемами (морем или рекой), так как они заполняются
всякий раз, как поднимается уровень воды в реке или на побережье.
(Белевич,1958).
Значительные площади по
окраинам ильменей покрыты новейшим аллювием, где растут влаголюбивые растения:
чакан (Typha angustifolia L.), тростник (Phragmites communis
Trin.) и др.
Степная часть более
однообразна по орографии и литологии. Местность преимущественно полупустынная,
с небольшими частыми понижениями, засоленными почвами и соответствующей
растительностью: шведки (Suaeda maritime
Dumort.), а в более засоленных впадинах —
солянки (Petfosimonia crassifolia Bge, Salicornia herbacea
L.) и др.
С севера на юг характер
рельефа также изменяется. Здесь можно выделить три участка: северный, средний,
или Долбанский, и южный, или Лаганский. (Белевич,1958).
Северный участок, до
ильменя Большой Чапчалган, характеризуется развитием относительно узких и
высоких бугров, расположенных правильными рядами с запада на восток и
разделенных ильменями. Ильмени глубоки и связаны с протоками Волги. Террасы
морфологически не выражены. В склонах бугров наблюдается асимметрия: крутые —
северные, отлогие — южные склоны. (Белевич,1958).
Средний или Долбанский участок
характеризуется развитием на территории нового вида ильменей, расчленяющих
поверхность Калмыцкой степи не только в направлении широтном, но и глубоко с
севера на юг. К таким ильменям относятся Б. Чапчалган, Харцаган, Дамбур и
Харбата. Соответственно этому и бэровские бугры вокруг указанных ильменей
занимают большие площади и имеют очертания, растянутые с севера на юг. Особого
внимания заслуживает ильмень Б. Чада. Очертания берега и литологический состав
приильменной толщи заставляют думать о происхождении этого ильменя из култука.
(Белевич,1958).
Другая часть бугров
сохраняет тот же характер, что и на севере. К югу бугры уплощаются, межбугровые
пространства делаются шире. Паводковые воды на уровень воды в ильменях
оказывают меньшее влияние. Возрастает значение морян. Увеличиваются площади,
занятые под камышовыми и чакановыми зарослями.
Наконец, южный, или Лаганский,
участок находится к югу от Долбана. Бэровские бугры следуют не с запада на
восток, а с некоторым отклонением, на восток-юго-восток. Расстояние между
буграми весьма значительно. Бугры широкие и низкие. Ильмени широкие, но
мелководные, ориентированы они, как и бугры, в сторону Каспийского моря. Подъем
воды на Волге в ильменях слабо отражается. Наполняются они водой
преимущественно со стороны открытого моря, поэтому уровень воды в них часто
колеблется в больших пределах, а при сильных сгонах часть ильменей совершенно
лишается воды. Вода солоноватая и горьковатая. В прошлом часть ильменей, открытых
к морю, была постоянными заливами - култуками. Бугры южного участка более,
густо заселены растительностью. Громадные прибрежные площади заняты
розоцветными солянками (Salsola rosacea
L.) с примесью луковника (Heleocharis sp.) и трехгранки (Scirpus maritimus L.), реже встречается угнетенный чакан
(Typha angustifolia L.).
В западной степной части основным и наиболее
резко выраженным элементом рельефа Калмыцких степей являются бугры. В основной
массе они представлены морфологически обособленными холмами — увалами,
ориентированными с запада на восток. Иногда встречаются целые массивы в виде плато.
На бугровых массивах, за счет разрушения верхней толщи из глинистого песка и
супеси, нередко происходит образование барханных песков или мелкобугристого
рельефа. (Белевич,1958).
Межбугровые понижения
выражены в виде частых и правильных по расположению депрессий волнистой степи.
Они чередуются с буграми и в поперечном профиле представляют мягкое понижение
или погнутую кривую с еле заметным более пологим северным склоном. Продольный
профиль их имеет вид ровной линии со слабым наклоном к открытым водоемам. Часть
межбугровых понижений или котонин заполнена солями. Развитие самосадочных озер
происходит, главным образом, в степной части западных ильменей; в восточной
части они выражены слабее. Кроме хлористых солей, образуются и важные в
промышленном отношении сернокислые: глауберова соль и астраханит.
(Белевич,1958).
Грязи сильно осолоненных
межбугровых понижений обладают высокими лечебными свойствами и используются для
этих целей (грязелечебница курорта Тинаки пользуется грязями соленого
близлежащего озера). Озеро расположено в глубокой и широкой межбугровой
котловине. Соль тонкой корочкой покрывает низкую окраинную часть котловины.
Средняя часть озера наполнена рапой — солевым раствором, откуда и выбирается
грязь. Выпоты солей широкой полосой покрывают склон и за пределами самой
котловины (до 20—30 см высоты). Растительность исключительно солончаковая —
солерос (Salicornia herbacea L.), розоцветная солянка (Salsola rosacea L.) и реже сарсазан (Halocnemum strobilacerum L.).
Эрозионные процессы на
описываемой территории обязаны почти исключительно действию волны на широких
просторах ильменей, подобно морской волне, подмывающей бугры. осыпи широкими и
низкими шлейфами отделяют бугры от ильменей. Они состоят из песков хорошо
отмытых обедненных окисными соединениями железа. По механическому составу
(соотношению отдельных фракций) пески близко подходят к бугровым, но в еще
большей степени лишены частиц <0,05 мм. (Белевич,1958).
Овраги не развиты,
глубокие овражки были встречены между ильменями Культкун и Мюн-Цаган по южной
окраине бугра. Они имели в длину до 4 —5 и 1—2м в глубину. Н. Г. Краснова в
работе, посвященной береговой полосе западных и восточных ильменей (1938 г.),
приводит описание оврагов на восточной оконечности бугра Буйна-Газыр, где на
протяжении 0,5 км находится 14 оврагов, из которых некоторые достигают 40 м в
длину и 3—3,5 м в глубину. (Брюшков, 1951).
Проявление эрозионных
процессов можно видеть в долинообразных неглубоких понижениях, лучеобразно
расходящихся по склонам от вершины бугра. Возможно, что началом для их
образования послужили проселочные дороги, часто идущие по буграм. Из дорожной
выбоины током дождевой воды песчаный материал был вынесен, поэтому и
образовалось понижение; второстепенные же борозды могли образоваться в период
избыточного количества дождевой воды. (Брюшков, 1951).
Эоловые процессы развиты
больше. Песчаные и супесчаные отложения степи в условиях сухого климата, бедной
растительности и сильного ветра легко переотлагаются и дают начало бугристым
весьма подвижным пескам. Особенно интенсивно эоловые процессы протекают в
западной части района, где ильмени осушились и толща малосвязанного иссушенного
песка передвигается вместе с ветром. В местах, заселенных кустарниковой
(тамарикс) растительностью, кочующий песок задерживается. Следует отметить, что
на направление дефляционных процессов большое влияние оказывает деятельность
человека, не всегда ясно представляющего себе тот огромный вред, который
получается вследствие разрушения растительного покрова (например, вследствие
долгого выпаса скота на одном месте).
На поверхности Калмыцкой
степи в удаленных от населенных пунктов местах встречаются участки, сплошь
покрытые земляными пирамидальными кучами, выброшенными сусликами. Размер этих
пирамидок 40-50 см. по высоте и 50- 60 см. по основанию, объем около 0,25м3
. Пески, слагающие их, также отчасти служат источником образования кочующих
бугровых песков. (Брюшков, 1951).
2.5.
Эколого-географическая характеристика ильменя Горчичный
Ильмень Горчичный
расположен в 35 км от г. Астрахань
и занимает площадь 200 гектаров. Имеет длину 4 километра и ширину до 600
метров. Этот водоем мелководен: максимальная глубина достигает 4,2 метра.
Глубина до 2 метров занимает почти 50 % площади ильменя. Ильмень изолирован от
других ильменей. Средняя зарастаемость 25 - 30 % от общей площади. Грунты илистые,
или песчано-илистые. По данным гидрохимического анализа воды 1999 года, средняя
соленость 1—4 г/л. Кислородный режим в пределах нормы (6,02-7,87).
Минерализация происходит, в основном, за счет солей хлоридно-натриевого
состава. Сезонная биомасса равна 2,5 — 30 г/м3.
Имеются зоны
сероводородного заражения. Ильмень Горчичный относится к трактовым ильменям
западной подстепной зоны Астраханской области(Отчет..., 1995).
Характерной особенностью
ил. Горчичный, как и большинства других ильменей, является огромное испарение
влаги с площади водяного зеркала, испарение превышает количество выпадающих
осадков примерено в 10 раз и поэтому, чтобы не нарушать гидрологический режим
ильменя, необходима постоянная его подпитка пресными водами. Подача воды в
ильмень была налажена АГТУ и прудовым комбинатом в конце мая 1994 года от
ильменя Чистая Шейна, при помощи специально прорытого канала. В связи с этим,
разные участки ильменя имеют разную соленость. На основании этого ильмень был
разделен на несколько основных станций, с которых и производится отбор проб
бентосных, планктонных организмов, а также воды на предмет выяснения ее
химического состава. Как указывалось выше, ильмень опресняется с помощью
искусственно прорытого канала, по которому поступает пресная вода, что,
позволяет успешно выращивать в ильмене рыбу, вторая естественно защищена от
некоторых видов паразитов соленой водой.
Вода имеет
удовлетворительные органолептические показатели (Отчет…, 1995).
Динамика подвижных форм в
донных отложениях определяется механическим составом грунтов, гидробиологическими
условиями водоема, кроме того, в значительной степени поглотительной
способностью донных осадков, прочностью связывания ими металлов,
окислительно-восстановительными условиями среды.
Усредненные данные по
содержанию металлов в грунтах изучаемого водоема свидетельствуют об их
вариабельности в отдельные месяцы:
Апрель - Zn> Mn > Ni
> Pb > Cu > Со > Cd;
Июль -Mn > Ni > Zn
> Cu > Pb > Со > Cd.
Данные по содержанию
металлов в грунте водоемов в отдельные месяцы представлены в таблице 14.
Таблица
14
Содержание металлов в грунте ильменя Горчичный в 2002 г.
| Месяц |
Cu |
Zn |
Mn |
Co |
Ni |
Pb |
Cd |
|
|
мг/кг |
| Апрель |
43,0 |
763,6 |
521,1 |
34,0 |
102,3 |
42,3 |
2,5 |
| Май |
9,5 |
1128,3 |
236,4 |
15,5 |
32,0 |
32,5 |
3,3 |
| Июль |
7,0 |
265,3 |
2,0 |
10,5 |
34,0 |
9,0 |
11,0 |
| Август |
19,57 |
49,0 |
201,3 |
14,5 |
25,3 |
16,0 |
0,5 |
| Среднее |
19,76 |
551,26 |
245,2 |
18,62 |
48,4 |
25,9 |
1,83 |
Отмечается факт
увеличения концентрации отдельных элементов в грунте в мае, то есть вовремя
паводка.
В начале лета концентрация
всех элементов снижается, а к осени – несколько возрастает. Наиболее отчетливо
эта закономерность проявляется в поверхностном слое грунта в пределах 0,5 — 5,0
см .(табл. 15).
Таблица
15
Концентрация металлов в
слоях донных осадков ил. Горчичный в августе 2002 г.
| Слой, см |
Cu |
Zn |
Mn |
Co |
Ni |
Pb |
Cd |
|
|
мг/кг |
| 0,5-1,0 |
9,32 |
52 |
86 |
3,56 |
7,6 |
6,58 |
0,11 |
| 2,0-3,0 |
19,3 |
48 |
141 |
11,5 |
16,3 |
17,29 |
0,3 |
| 4,0-5,0 |
19,3 |
52 |
145 |
15,64 |
17,5 |
21,64 |
0,2 |
| 6,0-7,0 |
20,5 |
56 |
157 |
15,68 |
33,6 |
19,76 |
0,5 |
| 8,0-9,0 |
23,4 |
40 |
265 |
16,55 |
46,5 |
20,31 |
0,8 |
| 9,0-10,0 |
25,6 |
46 |
302,5 |
24,45 |
30,4 |
10,71 |
0,7 |
| Среднее |
19,57 |
49,0 |
201,3 |
14,5 |
25,3 |
16,0 |
0,5 |
Обнаруженная
закономерность, скорее всего, связана с тем, что верхние слои донных осадков
подвержены различному влиянию, как со стороны воды, так и со стороны живых организмов,
поскольку отбор был произведен летом, то данные анализа отражают лишь
конкретную ситуацию, когда большое количество металлов из грунта было вовлечено
в трофический круговорот. (Горбунов, 1976)
Наблюдаемые колебания
химического состава окружающей среды имеют существенное значение в жизни
гидробионтов, у которых абсорбция минеральных веществ непосредственно из воды
занимает ведущее место в общем балансе (Отчет..., 1995).
Глава 3.
Материалы и методы
Материалом для настоящей
работы послужили результаты исследований с 1996 г. по 2003 г. в районе Западно-подстепных ильменей.
Для изучения был выбран ильмень Горчичный, который расположен В западной части
от реки Волга в сторону города Элиста, в 35 км от города Астрахани в районе
села Николаевка.
Пробы обрабатывались в
лаборатории кафедры «Гидробиологии и общей экологии».
3.1.
Методы сбора зоопланктона
Универсального метода
сбора зоопланктона, пригодного для всех групп организмов и для всех типов
водоемов, не существует.
Все известные методы
могут быть разделены на две группы: 1) отделение планктона от воды в самой
воде (планктонные сети, планктонные тралы, планктоночерпатели); 2)
зачерпывание или насасывание воды с отделением планктона от воды после подъема
прибора на поверхность путем фильтрации через сетку или отстаивания, а также
Центрифугирования.
А. Сетяной метод. Лов планктона сетями. Вода,
содержащая планктон, -фильтруется через специальную сеть из материала,
пропускающего воду и задерживающего планктон. Такими материалами являются
шелковое мельничное сито, капрон, найлон и др. Мельничное сито имеет различную
плотность, обозначаемую номерами от 7 до 77, каждый из которых соответствует
числу ячей в 10 мм ткани.
Самое редкое сито № 7
имеет размеры ячей 1,364 мм, а самое плотное № 77 - 0,064 мм. Мельничное сито -
материал довольно дорогой, не очень прочный, его не всегда легко получить.
Кроме того, с течением времени сито деформируется и размеры ячей, как правило,
уменьшаются. В настоящее время для изготовления сетей широко используют капрон
и найлон; они очень прочны, их ячеи имеют почти правильную квадратную форму, а
размер не изменяется и при долгом употреблении. Благодаря гладкости нитей
капрона и найлона организмы к ним не приклеиваются, тогда как в мельничном сите
они легко застревают. Толщина нитей капрона и найлона меньше, чем у мельничного
сита, и поэтому нумерация их разная. Например, № 38 мельничного сита
соответствует № 49; № 64 соответствует № 73.
Различают сети
качественные, предназначенные для массового сбора планктона, и количественные,
с помощью которых производится количественный учет организмов.
Качественные сети имеют
форму усеченного конуса, реже цилиндра, изготовленного из фильтрующего
материала. Широким концом мешок пришивают к металлическому (обычно латунному)
кольцу, а к узкому концу прикрепляют съемный металлический (или плексигласовый)
стаканчик различной конструкции. В нем концентрируется отфильтрованный из воды
планктон. Материал для сетного конуса раскраивают по выкройке с помощью
различных приемов. А. Липин рекомендует для небольшой модели качественной сети
(диаметром 25 см) взять 0,5 м ткани, отрезать от нее квадрат со стороной 50 см.
На нем карандашом проводят диагональ. Из концов ее радиусом, равным стороне
квадрата, проводят большие дуги АВ от двух других углов до пересечения с
диагональю и малые дуги радиусом 10 см. Затем квадрат разрезают по диагонали, а
каждый из получившихся треугольников — по дуге. Прежде чем сшивать треугольники
(половинки конуса), по той же выкройке вырезают из плотной ткани (бязь, холст и
др.) дуговые полоски а и б и нашивают на половинки конуса шириной
соответственно 5(а) и 2(6) см. Обрезав вершины обеих половинок конуса по самой
короткой дуге, сшивают обе половинки в целый конус, основанием которого
обшивают металлическое кольцо различного диаметра. К нему на равном расстоянии
друг от друга прикрепляют три прочные бечевки, свободные концы которых
связывают вместе над входным отверстием сетки или привязывают к небольшому
кольцу, к которому присоединяется трос для спуска сети. Вершину конуса прикрепляют
к планктонному стаканчику длиной 6—7 см, диаметром 4 см.
Для качественных сборов
пресноводного планктона употребляют разные модели сетей, причем чаще всего сеть
Апштейна длиной 55—100 см и диаметром 25—40 см.
В водоемах глубиной до
1,5 м используют сеть Липина с металлическим воронкообразным дном. Ею можно
ловить на расстоянии всего нескольких сантиметров от дна.
Для лова планктона на
течении или по ходу судна используют цилиндрические сетки. Для сбора планктона
у берега иногда зачерпывают воду каким-либо вымеренным сосудом с последующей
фильтрацией воды через небольшую сеть из плотного мельничного сита или капрона.
Для сборов морского планктона применяют сети больших размеров длиной до 1 м и
диаметром более 1 м.
Количественные сети на
переднем конце имеют надставку различной формы из плотной материи (парусина,
брезент, холст и др.) для ослабления обратных токов воды, образующихся при
протягивании сети. Благодаря надставке сети имеют два металлических кольца. При
конструировании количественных сетей размеры их рассчитывают так, чтобы
пропускаемая через сеть вода действительно фильтровалась. Для этого нужно,
чтобы сумма размеров всех ячей была больше площади входного отверстия сети.
Для вертикальных ловов
наиболее часто используют сеть Апштейна и сеть Джеди.
В пресных водоемах широко
применяют сеть Эпштейна с надставкой в форме усеченного конуса. Размеры Средней
'модели этой сети:
диаметр кольца входного отверстия 14 см, второго кольца 40 см, длина надставки
20 см, длина фильтрующей части 90-100 см
Замыкающаяся сеть Джеди
имеет высокий надставной усеченный конус, и поэтому второе кольцо находится
примерно посредине длины сетки. Сеть закрывается путем опрокидывания надставки.
Она сбрасывается с троса с помощью замыкающего механизма. По тросу посылается
специальный посыльный груз, который ударяет по замыкателю, передний конус сети
перегибается и закрывает входное отверстие. Сеть Джеди, отличающуюся хорошей
уловистостью, широко используют
как на пресных, так и на морских водоемах. Скорость подъема сети должна быть не
менее 0,25 и не более 0,5 м/с.
Для горизонтальных ловов
применяют замыкающиеся сети различных конструкций. Нередко фильтрующая сеть
заключена в металлический футляр со щелями, сквозь которые проходит вода,
выходящая через стенки сети. Входное отверстие сетей, предназначенных для
горизонтальных ловов, прикрепляется к массивной металлической раме и
обеспечивается системой посыльных грузов и замыкающих механизмов. Для учета
профильтрованной воды пользуются различными счетчиками.
Орудия лова планктона по
ходу судна. Основу этих орудий составляет планктонная сеть, заключенная в
корпус, чаще всего металлический. Входное отверстие сетей делают, возможно,
малым, а поверхность фильтрующего конуса возможно большей. Вследствие малой
площади входного отверстия не возникает большого давления на стенки сети, и она
не рвется.
Стандартный планктонный и
индикатор Гарди представляет собой цилиндрическую трубу длиной 56 см с
диаметром входного отверстия около 4 см. Задний конец прибора открывается, что
позволяет вставлять в трубу кольцо с диском из фильтрующего материала (обычно
мельничное сито № 23). Прибор снабжен системой пикирующих плоскостей на
переднем конце и плоскостями-стабилизаторами на заднем конце. Эти
приспособления позволяют индикатору быстро погружаться и плыть на глубине 8—10
м. Планктон концентрируется на шелковом диске, который периодически вынимают и
заменяют чистым. Уже при беглом осмотре его можно судить о степени развития
планктона и его составе. Более подробный анализ собранного планктона производится
в лаборатории. Первоначально индикатор Гарди был предназначен для изучения
горизонтального распределения планктона и установления связи между его
количеством, качеством и численностью рыбы. В настоящее время более надежные
указания на присутствие рыбы дает эхолотирование. Однако индикатор Гарди широко
применяется для изучения планктона.
Трал Айзеке—Кидда состоит
из конической сети, прикрепленной к большой U-образной распорной доске. Размеры входного отверстия трала
3—4,5 м. Трал снабжен приборами, регистрирующими объем профильтрованной воды и
глубину сбора. Работать тралом можно до глубины 3700 м. Особенно пригоден он
для лова макропланктона и мелкого нектона.
Помимо описанных приборов
сравнительно простого устройства применяется ряд автоматических собирателей,
которые могут производить непрерывные сборы на больших расстояниях.
Для своевременного сбора
планктона с разных глубин вдоль одного троса прикрепляют несколько сетей на
некотором расстоянии одна от другой.
Лов планктона
планктоночерпателями (планктонособирателями). Эти приборы работают по принципу
батометров, т. е. вырезают определенный объем воды вместе с содержащимся в нем
планктоном. Стенки прибора выполнены из газа, верхняя и нижняя крышки —
металлические. В приборах Богорова, Вовка и некоторых других крышки неподвижны.
Планктоночерпатели более совершенной конструкции имеют подвижные крышки,
которые при спуске прибора находятся в вертикальном положении. Этим уменьшается
сопротивление его окружающей воде. Кроме того, не отпугиваются организмы. Объем
планктонособирателей колеблется от 5—10 (пресные воды) до 25—50 л (морские
водоемы). Уловистость планктонособирателей в 2—5 раз выше, чем сетей.
Недостатком их, однако, является небольшой объем — несколько десятков литров.
Сетяной метод сбора
планктона широко распространен. Большим достоинством его является возможность
облавливать большие объемы воды (в десятки и сотни кубических метров).
Однако он отличается и
рядом серьезных недостатков. Источником ошибок является неточность учета объема
профильтрованной воды. Предполагается, что количество ее равно объему цилиндра,
основанием которого служит входное отверстие сети, а высотой — протяженность
лова, устанавливаемая тем или иным путем. Такой способ применим лишь при лове
планктона на небольшой глубине (2—5 м). При глубоководных сборах для учета
профильтрованной воды пользуются счетчиками различной конструкции, работающими
не всегда удовлетворительно, тем более что часть воды, несмотря на надставку
выбрасывается из сети, и учесть это количество трудно. Фильтрующая способность
сети с течением времени уменьшается, так как происходит деформация ячей сетного
материала (мельничный газ) и их засорение детритом и застревающими здесь
организмами. Существенным источником ошибок является малая скорость движения
сети. В результате представители мезо- и макропланктона активно уходят от
сетей. Еще один источник ошибок — неравномерное, пятнистое распределение
организмов в водоеме. В результате численность представителей планктона,
собранных даже в двух соседних пунктах, может различаться в сотни и тысячи раз.
Для уменьшения влияния на результаты исследования неравномерности
горизонтального и вертикального распределения планктона пользуются
репрезентативной (представительной) пробой. Ее получают путем смешения ряда
проб, взятых в разных местах и на разных глубинах.
Несмотря на существенные
недостатки сетяного метода лова, он используется весьма широко. Советские
исследователи считают, что в морских исследованиях наиболее применима сеть
Джеди, особенно ее большая модель длиной свыше 3 м. Высокая уловистость сети
Джеди определяется ее конструкцией — способом замыкания и отсутствием вымывания
организмов из сети в момент ее закрытия. Обычно сетяной метод лова планктона
применяют в сочетании с другими методами.
Б. Метод зачерпывают
воды. Для зачерпывания
воды используют приборы различного типа. Для изучения планктона в рыбоводных
прудах часто пользуются прибором Ляхновича. Он состоит из металлического
цилиндра длиной 120 см и диаметром 3 см с запирающимся приспособлением. Общая
вместимость прибора 6 л. Сверху к цилиндру припаяна крышка с отверстием, через
которое пропущен стержень с рукояткой сверху и запирающей нижней крышкой.
Прибор с лодки или плота погружают в открытом виде до дна, а затем опускают
цилиндр на нижнюю запирающую крышку и с помощью ручки отводят стержень в
боковой зазор верхней крышки. Поднятый на поверхность прибор открывается, и
вода процеживается через планктонную сеть.
Для сбора планктона на
озерах и водохранилищах часто используют планктонобатометр Дьяченко—Кожевникова
(ДК). Он состоит из укрепленного на стойке цилиндра вместимостью 10 л с
верхними и нижними крышками. На нижней крышке имеется кран. Перед началом
работы крышки закрепляют в вертикальном положении. В таком виде прибор опускают
на нужную глубину и резко останавливают. От толчка обе крышки закрываются.
После подъема прибора кран открывается и вода процеживается через сачок из газа
№ 70. В течение всей работы с планктонобатометром сачок хранят в воде, чтобы
его материал не деформировался. В СССР и за рубежом широко применяется батометр
Рутнера. Он имеет вид открытого с обоих концов цилиндра из стекла или
плексигласа диаметром 8—10 см и вместимостью 1 -1,5 л. Прибор опускают в водоем
в открытом виде. На нужной глубине сильным встряхиванием за трос или с помощью
посыльного груза, ударяющего по спусковому механизму, батометр на обоих концах
наглухо закрывается крышками.
При морских исследованиях
употребляют более сложные батометры. Батометр Стеман—Нильсена представляет
собой конический мешок из парусины, пропитанный олифой, длиной 1,25 м,
диаметром 0,5 м, вместимостью 100 л. Прибор опускают на нужную глубину в
сложенном виде. Там с помощью посыльного груза он раскрывается и наполняется
водой. Закрывание производится с помощью второго посыльного груза, который
освобождает замыкательный шнур, перетягивающий мешок в его верхней части.
3.2.
Методы обработки зоопланктона
1. Объемный метод. Для
измерения объемов планктона используют в основном способ вытеснения жидкости
или метод плотного объема. При этом определяют объем жидкости, вытесненной
планктоном, освобожденным от прилипшей и промежуточной воды. Этот метод дает
хорошие результаты при изучении проб, содержащих преимущественно зоопланктон;
при большом количестве фитопланктона из осадка трудно удалить промежуточную
влагу. Для измерения массы сырого вещества планктона используют мерный цилиндр,
бюретку или волюменометр.
В первом случае пробу
планктона отфильтровывают через шелковый газ и после легкого обсушивания
фильтровальной бумагой переносят на кусочке газа (объем которого во влажном
состоянии определяют заранее) в мерный сосуд (цилиндр или бюретка). Разница
между высотой жидкости в измерительном сосуде до и после погружения в нее
планктона дает величину плотного объема измеряемой пробы.
Волюменометр Яшнова состоит
из стеклянного цилиндра, открытого с обоих концов и имеющего небольшой бортик
на нижнем конце; металлической крышки с двумя отверстиями, в одно из них
пропускается тонкий, заостренный снизу стержень, закрепляемый в желаемом
положении винтом; кружка пластинчатой резины для закрытия цилиндра снизу, а
также кружка мелкоячеистого сита. Волюменометр имеет разные размеры: малая
модель вместительностью около 80 мл, высотой 8 см, с внутренним диаметром 3,6
см; большая модель имеет размеры соответственно 300 мл, 15 см и 5 см. Перед
началом работы прибор тарируют. Для этого на нижний конец цилиндра туго
натягивают кружок пластинчатой резины. Этим обеспечивается по крайней мере 10
мин герметичность замыкания цилиндра. Из бюретки в цилиндр вливают 50 (для малой
модели) или 200 мл (для большой модели) воды, и штифт опускают до
соприкосновения его острия с поверхностью жидкости в цилиндре. Волюменометр
готов к работе. воду, налитую для тарирования, удаляют и в волюменометр вливают
пробу планктона. Затем снимают резину, воду отфильтровывают через сито и
цилиндр. Для удаления излишней влаги его ставят на фильтровальную бумагу. Затем
снова закрывают резиной. Из бюретки наливают раствор 4% - ного формалина до тех
пор, пока острие шрифта не коснется поверхности жидкости. Разница в показаниях
бюретки по сравнению с влитой в цилиндр при тарировании водой служит мерой
вытесненного объема планктона. Объемный метод удобен в экспедиционных условиях.
К недостаткам метода следует отнести сложность удаления промежуточной воды, так
как при этом возможно обезвоживание самих организмов.
3.3.
Методы сбора фитопланктона
3.3.1.
Методы сбора и орудия лова фитопланктона
В последнее время
гидробиологами довольно редко используется метод сетяного лова, который
предназначен для отбора качественных проб фитопланктона всех категорий, кроме
наннопланктона. Однако в некоторых случаях он остается наиболее эффективным,
особенно когда тот или иной вид представлен в незначительном количестве и может
быть собран только в результате процеживания большого объема воды. Для
выявления видового состава фитопланктона лучше использовать планктонную сеть
Джеди, изготовленную из очень мелкого (№70 и ещё больших размеров) мельничного
сита шёлковой или капроновой нити. Материал, отобранный сетью, может быть просмотрен
в живом состояние в полевых условиях.
Одинаково применим для
качественного и количественного сбора батометрический метод отбора проб фитопланктона.
Системы существующих батометров весьма разнообразны. Почти все конструкции и их
описания приведены в монографии И.А. Киселёва. Опыт работы показал, что
батометры типа батометра Руттнера малопригодны для сбора проб фитопланктона,
так как, погружаясь в водоём, они своим нижним диском разбивают поверхностную
плёнку и перемешивают организмы водной толщи в районе действия прибора.. Необходимо
пользоваться приборами, у которых при погружении обе створки находятся в
вертикальном положении и не мешают вырезанию определённого столба воды.
Наиболее прост в
изготовлении и удобен в работе батометр А.В. Францева. Его способность вырезать
метровый слой жидкости особенно ценна при исследовании вертикального распределения
водорослей. При комплексных работах, когда необходимо получить одновременно
воду для биологического и химического анализов, следует применять батометры большего
рабочего объёма. К таким приборам относится планктобагометр ДК (Дьяченко -
Кожевниковой), ёмкость которого у большой модели равна 10, а у малой - 5л. Ещё
более удобен батометр Молчанова ГР-18. Он предназначен для взятия проб воды с
различных глубин водоема
и одновременного измерения температуры воды исследуемого слоя (от 1 до 40 °С).
Батометр ГР-18 имеет два цилиндра из органического стекла, емкость которых не
менее 4л.
В быстротекущих водах
отбор проб этими приборами осложнён из-за эффекта сноса. Для таких водоёмов
применяются батометры Жуковского или Фридингера, а также батометр
сконструированный Яагом, Амбюлем и Циммерманом. В воду батометры указанных
конструкций опускаются с открытыми горизонтально крышками; вода при этом
свободно проходит насквозь.
3.4. Методы обработки фитопланктона
3.4.1.Методы
сгущения и консервации фитопланктона
Наиболее
распространёнными методами консервирования планктона являются седиментация и
фильтрация пробы воды через мелкопористые мембранные фильтры. Седиментационный
(осадочный, или остаточный) метод, предложенный Р.Г. Гринбергом ещё в 1915г. и
модифицированный П.И. Усачёвым, распространён и в настоящее время. Метод
заключается в отстаивании законсервированной исследуемой пробы воды в тёмном
прохладном месте. Объём пробы зависит от степени развития фитопланктона; обычно
он составляет 0,5л, а для олиготрофных водоёмов — 1л. Исследователи предлагают
разные сроки отстаивания пробы. По нашему мнению, следует отстаивать пробу не
менее 10 дней.
Для фиксации проб
отдельными гидробиологами до сих пор применяется формалин, но он разрушает
нежные флагеллаты и не ликвидирует газовые вакуоли у синезелёных, что мешает их
осаждению. Уже в 1926г. Усачёвым было предложено перейти на фиксацию проб
йодистым калием. Позднее Утермель рекомендовал рецепт ещё одного фиксатора,
трёх капель которого вполне достаточно для фиксации 100мл планктонной пробы. На
основании этого раствора в Институте биологии внутренних проб РАН разработан
фиксатор, состоящий из двух растворов:
Раствор I Раствор II Раствор III
Kl -10г Хромовая
кислота 1%-ная - 5см3
H2O -50см3 Ледяная
уксусная кислота - 10 см3
I -5г Формалин
40%-ный - 80см3
Оба раствора сливаются и
хранятся в тёмном месте. При применении йодных фиксаторов в клетках водорослей
хорошо обнаруживаются пиреноиды, жгутики, окрашивается слизь, исчезают вакуоли
у большинства имеющих их синезелёных. Наличие формалина в составе консерванта,
позволяет хранить пробу длительное время. Фиксированная проба после отстаивания
концентрируется отсасыванием воды с помощью трубки-сифона с загнутым на 2см
вверх концом, затянутым газом №№ 70-76 или с помощью устройства для
автоматического консервирования фитопланктонных проб, предложенного А.Ф.
Крахмальным. Устройство состоит из двух расположенных на разных уровнях
штативов (на верхний штатив устанавливается сосуд с концентрируемой водой, на
нижний — мерный цилиндр, в который отсасывается вода), сифона, трубок, двух
вентилей (обычного и соленоидного) и следующего устройства. Это устройство
позволяет создать стандартные условия концентрирования, а также отрегулировать
скорость отсасывания воды таким образом, чтобы исключить возможность попадания
в фильтрат мелких видов фитопланктона.
После отсасывания остаток
пробы в 30 -80 мл переливают в склянку (типа аптечной плевательницы). Туда же
сливают воду после ополаскивания стенок сосуда, в котором происходило
осаждение.
Широкое применение в
гидробиологии получил метод мембранной фильтрации, который способствует быстрой
концентрации проб и даёт возможность просматривать фитопланктон в живом
состоянии. Отечественное производство мембранных фильтров было начато в 1931 –
1932 гг. экспериментальной фабрикой ультрафильтров Министерства коммунального
хозяйства РСФСР (Мытищи). Для сгущения фитопланктона пригодны фильтры №6 и №5 с
диаметром пор 2-5 мкм и 1,2 мкм соответственно. Фильтр №6 рекомендуется
использовать как предварительный при обильных пробах для ускорения процесса
фильтрации, а также для разделения крупноразмерной и мелкоразмерной фракции
фитопланктона. После фильтрации на фильтре №6 полученный фильтрат, содержащий
вторую фракцию, следует пропустить повторно через фильтр №5.
В 80-х годах был налажен
выпуск мембранных фильтров «Владипор» Казанским производственным объединением
«Тасма» им. В.В. Куйбышева, из которых для концентрирования фитопланктона
пригоден фильтр «10 с диаметром пор около 1мкм. В упаковках имеются
фильтры-подложки войлочного типа, на которые укладывается сам мембранный
фильтр, что предотвращает его разрыв и способствует равномерному распределению
осадка. В нашей стране получили распространение также чешские фильтры марки
«Сынпор-2» с диаметром пор 1,2мкм.
Сухие фильтры содержат в
своих порах воздух, который закупоривает их и затрудняет фильтрацию. Для
удаления воздуха фильтры нужно прокипятить в дистиллированной воде в течение
20-ЗОмин. Воду следует нагревать медленно, а кипячение должно быть спокойным,
так как при бурном нагревании и кипячении фильтры скручиваются и становятся
непригодными к употреблению. Кроме того, для удаления воздуха из пор фильтров
можно рекомендовать длительное содержание фильтров в дистиллированной воде
(перед помещением их на такое хранение фильтры необходимо несколько раз промыть
в дистиллированной воде).
Фильтрацию проводят под
вакуумом в воронке с пористым или сетчатым дном, на которое укладывают
мембранный фильтр. Воронку укрепляют на колбе Бунзена, которую через верхний
тубус шлангом соединяют с вакуумным насосом. Возможно соединение нескольких
воронок одной трубкой или системой гибких шлангов, что позволяет фильтровать
сразу несколько проб.
И.М.Балонов предложил
портативный прибор, очень удобный в экспедиционных условиях, где колба Бунзена
заменена дюралевым стаканом, в котором при транспортировке переворачивается и
фиксируется модифицированная фильтрационная воронка из органического стекла.
Для создания вакуума он использует насос от мотороллера или велосипеда. Масса
прибора вместе с насосом 1260 г, а размеры в собранном виде 233*94
мм.
Пробу фильтруют до
определенного объема, оставляя над фильтром столбик воды высотой 1 см, или до
момента, когда воды над осадком уже нет, но фильтр еще остается влажным. Затем
планктон осторожно смывают с фильтра мягкой кисточкой и просчитывают в счетной
камере. желательно сразу после фильтрации просмотреть живой материал, что
позволяет не только обнаружить новые формы водорослей, но и живую пробу, фильтр
помещают в пенициллиновую склянку объемом 20 мл, заливают 5-10 мл фильтрата и
консервируют до слабо – желтого цвета. В этом случае за 30 мин до фильтрации
можно провести предварительную консервацию пробы несколькими каплями фиксатора,
что предотвратит деформацию проб на фильтре, которая может иметь место при
фильтрации живой пробы.
Оценка точности
осадочного и фильтрационного методов, проведенная К.А. Гусевой, показала, что
довольно близкие результаты с отстойным концентрированием получаются только в
случае двойной фильтрации пробы. Причина состоит в том, что при обильных пробах
только такая двойная фильтрация обеспечивает равномерное распределение
отфильтрованных водорослей по площади фильтра. при одноразовой фильтрации
происходит сбивание организмов фитопланктона в кучи или даже склеивание их на
фильтре. поэтому результаты подсчета фитопланктона непосредственно на таких
фильтрах (особенно при большом увеличении микроскопа) обычно выше результатов,
полученных с помощью отстойного метода.
Несмотря на определенные
достоинства метода мембранной фильтрации (Это прежде всего возможность анализа
живого материала, а также быстрота сгущения проб при малом исходном объеме)
многие гидробиологи предпочитают использовать отстойный метод как более простой
и не требующий специальных установок.
Изучать организмы в живом
состоянии можно и в случае применения метода центрифугирования, который
позволяет быстро осадить водоросли. Однако применять его при количественном
учете фитопланктона не следует, так как центрифуга не осаждает синезеленые
водоросли, содержащие вакуоли, и организмы с меньшей плотностью чем вода.
3.4.2.
Этикетирование проб
Каждая проба снабжается
этикеткой, на которой указывают название водного объекта, номер станции,
глубину, орудие лова, дату сбора. Этикетка пишется на пергаментной бумаге
твердым карандашом или шариковой ручкой и вкладывается под прокладку крышки.
Для этикеток удобно использовать лейкопластырь, кусочки которого наклеивают на
банку или крышку, а затем подписывают мягким карандашом или ручкой. Иногда на
этикетке ставится просто номер, который соответствует номеру записанному в
журнале или полевом дневнике. В дневник вносятся дополнительные сведения о
погоде, температуре, цветности, прозрачности воды, глубине станции, визуальные
наблюдения о качестве воды и т.д.
3.4.3.
Камерная
обработка фитопланктона
Метод прямого
микроскопирования является самым трудоёмким, но пока единственным методом,
позволяющим точно идентифицировать виды, получить их размерные характеристики,
определить физиологическое состояние и подсчитать численность. Определение
качественного состава фитопланктона следует проводить до вида по наиболее
широко применяемым определителям. Кроме того, нужно учитывать новые данные по
таксономии и систематике, публикуемые в специальной литературе, в частности в
ежегоднике «Новости систематики низших растений», «Ботаническом журнале». При
этом всегда необходимо указывать источник, по которому проведено определение
вида.
3.4.4.
Методы
подсчёта водорослей планктона
Для количественной
обработки фитопланктона удобны счетные камеры «Учинская» или «Нажотга» объёмом
0,01; 0,02 и 0,05 см3. Процесс подсчета очень трудоёмок и требует
большой тщательности. Существенным моментом является наполнение камеры, перед
которым проба тщательно перемешивается продуванием воздуха через капилляр с
входным отверстием не менее 2 мм. Этим же капилляром вносится одна - две капли
фильтрата, и камеру быстро закрывают покровным стеклом. Пробе дают осесть в течение
нескольких минут.
Второй важный момент -
это количество просчитанных полос. К.А.Гусева считает, что в камере объёмом
0,05 мл при количестве водорослей несколько сотен и десятков тысяч в 1 мл можно
ограничиться просчётом двух полос из 40 имеющихся в ней, при нескольких тысячах
клеток в 1 мл необходимо просчитать всю камеру. В камере же объёмом 0,01 мл
только при количестве нескольких сот тысяч можно просчитать две полосы, при
нескольких десятках тысяч - пять полос и при нескольких тысячах - всю камеру. Г.В.Кузьмин
советует просчитывать каждую пятую полосу указанных камер, а при высокой
численности - каждую десятую. Определение численности водорослей лучше
проводить в камерах разных объёмов. Так, крупные и колониальные формы планктона
просчитывают в камерах большего размера (не менее 0,05 — 0,1 см3),
для остальных видов подходят и более мелкие (0,01 и 0,02 см3).
За счётную единицу
следует принимать клетку. Пересчёт общей численности производится по формуле:
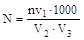
Где N - число клеток в 1
л воды исследуемого водного объекта; n - число клеток, обнаруженных в просчитанных полосах камеры; v1 - объём концентрата пробы, см3; V2 - объём воды в просчитанных полосах камеры, см3; V3 - объём профильтрованной пробы, см3.
3.5.
Методы
сбора бентоса
Орудия, применяемые при
изучении макрозообентоса (размером более 2 мм), подразделяются на орудия для
качественного и количественного сбора его.
Первые служат для
установления видового состава донной фауны. К ним относятся сачки, скребки,
драги, тралы.
Сачки состоят из
металлического обруча круглой или треугольной формы диаметром 20—30 см. Мешок
сачка выполняют из прочного материала (мешковины). Обруч сачка насаживают на
палку длиной 2—3 м. Сачок используют для сбора фауны зарослей, которая
представлена главным образом брюхоногими моллюсками, насекомыми и их личинками.
Скребок представляет
собой мешок из редкого прочного сита или мешковины, прикрепляемый к
металлическому ободу, который насаживают на палку длиной 2—3 м. Противоположный
край обода представляет собой заточенную с наружного края пластину длиной 6—18
см. Высота обода 8-20 см. Скребок служит для сбора фауны жестких грунтов. Драги
состоят из следующих частей: 1) мешок из прочного, редкого материала
(мешковина, частая дель, проволочные сетки); 2) металлическая массивная рама
различной формы. Рама закидной, драги треугольная; драга с ножами имеет
четырехугольную раму, верхний и нижние края которой затачивают.
Драгу с зубьями (зубчатую
драгу) применяют для сбора крупных двустворчатых моллюсков. Нижний и верхний
края ее снабжены длинными отогнутыми кнаружи зубьями. Мешок этой драги
выполняют из проволочной сетки. Овальная драга имеет раму овальной формы,
заточенную по всему наружному краю. Драга не застревает на камнях и срезает
прикрепленные к ним организмы. Размеры драг различны: обычно длина сторон рамы
равна 25—40 см, высота 20—35 см. Драги предназначены как для сбора организмов,
находящихся на поверхности дна (на камнях и другом субстрате), так и для
захвата грунта.
Тралы отличаются от драг
тем, что не захватывают грунт, а облавливают его поверхность, а также придонный
слой воды. В основе одного из них — бимтрале — лежат два так называемых дуговых
башмака, затянутых делью. Снизу и сзади между башмаками с большой слабиной
протянута цепь. Вверху башмаки соединены прочным деревянным брусом, называемым
бимом. К цепи и биму, а также к верхней части башмаков прикрепляют мотню,
снабженную иногда внутренним конусом. Верхняя части мотни несколько заходит
вперед и закрывает подвижную фауну, встречаемую тралом. Мотню выполняют из
частой дели или мешковины и защищают брезентовым фартуком. Размеры бимтрала:
длина башмака 40 см, его высота 25 см, длина епи 80 см, длина бима 70 см, длина
мотни 150 см. К качественным орудиям сбора относятся также камнедоставатели
разных систем. Их опускают на дно на шесте или на тросе в открытом состоянии;
лапы прибора охватывают камень и извлекают из воды.
3.5.1.Орудия
для количественного исследования зообентоса
Для количественных сборов
зообентоса можно в случае необходимости использовать качественные орудия -
сачки, скребки и др. Например, сачки применяют для количественного учета
личинок и куколок малярийного комара. При этом сачком диаметром 20 см проводят
у побережья 5 раз на протяжении 1 м и таким способом получают данные о
количестве личинок на площади 1 м2. Для получения количественных
проб с помощью скребка его погружают на заданную глубину и проводят по дну на
определенное расстояние. Умножая длину стальной пластины обода на длину полосы
облова, получают обловленную скребком площадь. Для количественных сборов
использовать можно также драги и тралы. Для этой цели к орудию прикрепляют так
называемый дрейфограф — прибор, который показывает скорость движения драги или
трала и пройденный ими путь.
Сконструированы
и специальные количественные драги, весьма точно учитывающие донную фауну,
однако довольно сложные. Некоторые количественные драги снабжены массивной
металлической рамой с ножом и полозьями. К раме прикрепляют мешок из
мелкоячеистой дели или другого прочного материала. Драгу закрывают крышкой; с
замыкающим механизмом. Благодаря полозьям такие драги скользят по дну, не
врезаясь в грунт. В зависимости от размеров рамы масса количественных драг
колеблется от 4—12 до 40 кг. Эксплуатируют драги с судна.
Специальными орудиями для
количественных сборов зообентоса служат дночерпатели разнообразной конструкции.
Схематически все дночерпатели можно разделить на две группы — штанговые и
тросовые.
Штанговые дночерпатели
имеют прямоугольную или цилиндрическую форму. У всех таких орудий в верхней
части имеются приспособления — держатели для штанги (шеста) из дюралевой трубки
или из дерева. Площадь захвата штанговых дночерпателей различна. Их используют
на небольшой глубине на относительно плотном грунте (песчаном или глинистом).
Дночерпатель опускают в
воду и погружают в грунт. С помощью различных приспособлений он замыкается и с
захваченным монолитом поднимается на борт судна. Здесь под дночерпатель
подводят кювету или таз, в которые выпадает захваченный грунт. Площадь захвата
приборов этого типа 0,01 м2. Штанговые дночерпатели применяют лишь
на глубинах, не превышающих длину штанги. На больших глубинах используют
тросовые дночерпатели различной формы. На мягких илистых грунтах применяют
коробочные дночерпатели, например типа Экмана—Берджа, с площадью захвата 1/25 и
1/40 м2. Этот прибор с сильными спиральными пружинами легко
приводится в действие простым поднятием рукоятки, поднимающей щеки прибора.
Срабатывает дночерпатель с помощью посыльного груза, ударяющего по спусковому
механизму, который освобождает пружины.
На очень мягких и
глубоких илах рекомендуется пользоваться дночерпателем Боруцкого с высоким
коробом, массой не менее 6 кг, чтобы обеспечить достаточно глубокое погружение
прибора в грунт.
Ковшовые дночерпатели в
виде нескольких моделей, с площадью захвата 1/10, 1/40 и 1/100 см2
имеют два изогнутых глубоких ковша, вращающихся на скрепляющей их оси. Тросиком
ковши соединяются с замыкающим приспособлением. При работе на больших глубинах
для утяжеления прибора к верхней части ковшей прикрепляют чугунные или
свинцовые пластины.
Наиболее совершенной
моделью ковшового дночерпателя является прибор "Океан-50". Его ковши
в верхней части имеют стальные крышки, которые свободно откидываются вверх, не
оказывая сопротивления при спуске прибора. Количество взятых на каждой станции
проб бывает различным и зависит от состава и количества бентоса, качества
грунта, а также от площади захвата прибором грунта. Рекомендуется при работе с
дночерпателями с площадью захвата 1/25 м2 брать не менее двух
выемок, а при меньшей площади (штанговые дночерпатели) — не менее 4—5 выемок.
3.6. Методы
обработки бентоса
Выборка и фиксация
материала. Пробы
бентоса, полученные описанными в теме 6 орудиями, отмывают от избытка грунта
различными способами.
Для отделения организмов
от мягкого грунта (ил, глинистый ил) пробы переносят в промывательное сито из
мельничного или капронового газа № 19—23, укрепленное в станке на борту или за
бортом судна. Сито, пришитое к четырехугольной металлической рамке, на веревке
можно опускать за борт на половину глубины мешка. При большом объеме пробы
промывку сначала проводят в станке, а затем за бортом судна. При работе с лодки
пользуются также вертикальным ситом Липина цилиндрической формы со стенками из
металлической сетки (с ячеёй 0,5 мм). Если проба содержит песчаный грунт, ее
переносят в таз и отмучивают. Песок взбалтывают рукой или палкой и воду со
взвесью многократно сливают в промывательный сачок из газа № 34—38. Затем
оставшийся грунт просматривают и находящиеся в нем организмы выбирают пинцетом.
Отмытые от избытка грунты
пробы в том случае, если не предусмотрена немедленная их разборка, помещают в
матерчатые мешочки или банки (стеклянные, пластмассовые), заливают 10 %- ным
раствором формалина, нейтрализованным содой (для получения 10%-ного формалина в
40%-ный его раствор добавляют воду из расчета 1:3), и снабжают пергаментными
этикетками, на которых указывают название водоема, № станции, дату, глубину,
орудие сбора. Однако разборку материала из отмытых проб рекомендуется
производить в полевых условиях до фиксации, так как живые организмы более
заметны и легче поддаются выборке.
Материал помещают
небольшими порциями в плоские сосуды (эмалированные ванночки, пластмассовые
кюветы и т. д.) и с помощью пинцета выбирают организмы. При обильном бентосе,
содержащем много мелких форм, можно пользоваться методом флотации (всплывания):
пробу по частям помещают в насыщенный раствор поваренной соли. Все организмы,
кроме моллюсков и олигохет, запутавшихся в растительных остатках, всплывают на
поверхность, и их выбирают маленьким сачком из газа. После этого грунт
просматривают под бинокуляром или с лупой. Организмы, собранные таким способом,
отмывают от соли. Организмы, обнаруженные в пробе, распределяют по крупным
систематическим группам (олигохеты, моллюски, ракообразные, личинки хирономид,
личинки ручейников и др.), помещают в пробирки, склянки или бюксы, снабжают
этикетками и фиксируют 10 % - ным формалином.
3.7.
Статистические исследования
Статистическая обработка
полученного цифрового материала проводилась с использованием программного
продукта Microsoft Excel XP.
Глава 4.
Результаты и их обсуждения
Исследование
фитопланктона ильменя Горчичного
За исследованный период «1996
- 1997, 2002 - 2003 гг. » было обнаружено 60 видов, из них 25 диатомовых, 21
зеленых, 9 сине-зеленых и 5 видов прочих организмов. Летом в фитопланктоне за
1996 и 1997 годы основа биомассы составили соответственно диатомовые (25, 967
мг/м3) и сине-зеленые (3085,746 мг/м3), а за 2002 и 2003
годы соответственно – зеленые (2588,714 мг/м3) и диатомовые (20,442
мг/м3) из общей биомассы за 4 года: 18111,846 мг/м3.
Основными доминирующими
видами в 1996 году из диатомовых были : Navikula sp., Nitzschia sp.; из
синезеленых - Microcustis sp., Gleocaspa.
В 1997 году из диатомовых
: Navicula sp., Pinularia sp., Cocconeis sp.,
из зеленых: Cosmarium sp.(табл. )
В 2002 году из диатомовых:
Navicula sp, Melosira
curvata, Nitzsnia linearis;
из зеленых: Chroococeum sp.
В 2003 году из
диатомовых: Navicula sp., Nitzschia acicularis.
из зеленых: Ulotrix voriosilis, Oocustis borgei,
из сине- зеленых: Gleocapsa minuta, G.
Turgida.
Распределение
фитопланктонов по годам неоднородно. Небольшим количеством видов
характерезуется 1996 год (около 11 видов), большая встречаемость отмечена в
2002 году (около 17 видов).
Анализируя полученные
данные можно сделать вывод о том, что Фитопланктон ильменя Горчичного
достаточно разнообразен по количественному и качественному составу, является
хорошей кормовой базой для фитопланктонафагов и беспозвоночных.
Исследование
зоопланктона ильменя Горчичного
За исследованный период
(1996, 1997, 2002, 2003 г. г.) было встречено 42 вида. Из них 11 ветвистоусых,
14 веслоногих, 14 коловраток и три прочих вида организма.
Летом в зоопланктоне за
1996 и 1997 г. г. основа биомассы составили соответственно ветвистоусые ( 230,
5 мг/м ) и коловратки (667, 74 мг/м ) , а за 2002, 2003 г. г. соответственно - ветвистоусые
( 696, 01 мг/м ) и ветвистоусые ( 2968, 503 мг/м ). Из общей массы за 4 года:
3960, 513 мг/м.
Основными доминирующими
видами в 1996 году были :
Из ветвистоусых : Bosmina longirostris,
Из веслоногих : Copepoditi, Nauplii,
Из коловраток: Brachionus guadridentatus, Keratella guadrota;
В 1997 году были :
Из ветвистоусых : Daphnia pulex,
Из веслоногих : Nauplius, Copepoditi,
Из коловраток: Brachionus guadridentatus, B. Urceus;
В 2002 году были :
Из ветвистоусых : Moina recnirostris, Chudorus biovatis.,
Из веслоногих : Cuclopoida, Harpastisoida;
В 2003 году были :
Из ветвистоусых : Moina rectirostris, Ceriodaphnia reticulata.,
Из веслоногих : Copepoditi, Nauplii,
Из коловраток: Brachionus guadridentatus,
Исходя из данных таблицы
видно, что количество зоопланктонов начиная с 1996 года по 2003 год увеличивается,
но большинство видов наблюдается в 2002 году.
Анализ структуру
зоопланктона ильменя Горчичного показывает, что доминирующее положение занимает
мелкие формы организмов ( веслоногие рачки ). По многочисленным литературным
данным эти организмы являются ( в виду их малого размера ) кормом в основном
для молоди рыб на ранних этапах развития.
4.3.
Исследование зообентоса ильменя Горчичный
Состояние зообентоса в
ильмене Горчичном в летний период за 1999г 2002, 2003 г.г. За этот период было
отмечено 23 вида, из них Mollusca 6
видов, Crustacea 6 видов, Insecta 11 видов.
Основную биомассу
зообентоса за1999г. составили Insecta
(146.2г/м ) Crustacea ( 1.48г/м) , Mollusca (48.66г/м). За 2002г.- Insecta(10.92 г/м), Crustacea (7.8 г/м) и Mollusca (3.6г/м). За 2003г.- Insecta(22.792 г/м), Crustacta (15.5г/м) и Mollusca(18.96г/м). Из общей биомассы за 3
года (265,721г/м).
Основными доминирующими
видами в1999году изInsecta были Chironomus albidus, из Mollusca- Gastropoda, а из Crustacea- Amphipoda. В 2002 году из Insecta были Chironomus albidus, а из Crustacea были Niphargoideus robustoides, а в 2003 году из Mollusca – Notonecta glauca.
Общая характеристика
распределения зообентоса в ильмене представлено в таблицах.
Данные таблицы
свидетельствуют о том, что общая биомасса зообентоса за 1999,год составила
196,343 г/м , а численность – 3680 экз/м . В 2002 году биомасса
составила-22,332 г/м, численность -9072 экз/м.
В 2003году биомасса составила
47,049 г/м , численность 5312,4 экз/м.
Заключение
Рыбоводство в солоноватоводных
ильменях – одно из перспективных направлений развития трудового хозяйства в
Астраханской области, что связано с большими площадями подстепных ильменей, их
достачточно высокой естественной биопродуктивностью. Кроме того, ильмени
представляют собой в большей своей массе изолированные водоемы что естественно
положительно сказывается на возможности управляемого развития их экосистемы в
нужную для человека сторону.
Неблагоприятными
факторами, связанными с возможностью использования подстепных ильменей
Астраханской области в качестве рыбоводных прудов заключается, прежде всего, с
тем, что они имеют нестабильный гидрохимический режим , что связано с их
мелководностью, большой зависимостью от поступления паводковых вод, что в
современных условиях зарегулированного речного стока представляет собой большую
проблему. Кроме того, препятствием на пути возможного использования этих
ильменей является климат Астраханской области, который характеризуется сильными
перепадами температур ( с особенно сильными положительными максимумами ), что
неблагоприятно сказывается на биоте обитающей в этих ильменях, так как
небольшие по объему и мелкие ильмени не обладают достаточной буферной
способностью. В связи с нерегулярной подачей воды, существует постоянная
опасность сильного засолонения ильменей, а большинство разводимых рыб
поддерживают нормальную жизнедеятельность в приделах солености равной 5 – 6 % ,
а соленость ильменей, не имеющих подпитки пресной водой может достигать 35 %. Плюсом
в возможности развития прудового хозяйства, и в частности выращивания белуги,
является то, что ильмени обладают достаточно большими запасами органического
вещества, необходимого для развития начальных звеньев пищевых цепей.
В ильмене Горчичном, как
и в любой другой гидроэко системе основой биопродуктивности, то есть
возможности для развития кормовых объектов является фитопланктон.
Фитопланктон ильменя
Горчичный представлен в основном тремя группами водорослей: сине-зеленые,
диатомовые и зеленые.
За исследованный период (
1996, 1997, 2002, 2003 г.г. ) было обнаружено 60 видов водорослей, из них
диатомовых – 25, зеленых – 21, сине-зеленых – 9 и 5 прочих организмов.
Можно сделать вывод о
том, что фитопланктоны ильменя достаточно разнообразен по количественному и
качественному составу и является хорошей кормовой базой фитопланктофагов и
беспозвоночных.
Следующим исследуемым
звеном, составляющим основу биопродуктивности ильменя, является зоопланктон.
В зоопланктоне за четырех
летний период встречено 42 вида из которых наибольшим числом характеризовались
веслоногие рачки. Среди этой группы организмов доминировали науплии и копеподии
, а также дафнии и Moina rectirostris. Коловратки представлены в основном
такими видами как Brachionus sp. и Keratella sp.
Сравнивая общее число
видов зоопланктона за 1996, 1997, 2002 и 2003 г.г. видно, что максимального
разнообразия зоопланктон достигал в 2003 году.
Анализ структуры
зоопланктона показывает, что в зоопланктоне ильменя Горчичного доминирующее
положение занимают мелкие формы ( веслоногие рачки ). По многочисленным
литературным данным эти группы являются ( в виду их малого размера ) кормом в
основном для молоди на ранних этапах развития.
Немаловажным элементом в
кормовой базе для подрастающей молоди являются бентические организмы. Бентос
является аккомулятором не только органического вещества но и большинство
вредных химических соединений, то есть является своеобразной буферной системой
в водоемах.
Общая биомасса зообентоса
за период с 1999г. по 2003г составляет 265.721г/м., а численность – 18053.3
экз/м.
Бентос ильменя
представлен, в основном, потребляемым как молодью, так и взрослой рыбой
формами, что является важной предпосылкой возможного вселения сюда рыб –
бентофагов.
По уровню
биопродуктивности зообентоса ильмень относится к низкопродуктивным
Планктоны и бентоносные
организмы являются основой для питания мелких видов рыб, которые в свою очередь
составляют основу пищевого рациона молоди белуги.
Выводы
Проведенные исследования
позволяют сделать следующие выводы.
1.
Фитопланктон
водоема достаточно разнообразен по качественному и количественному составу и
является хорошей кормовой базой для рыб фитопланктофагов и беспозвоночных.
2.
В зоопланктоне
ильменя доминирующее положение занимают мелкие формы веслоногих рачков, которые
являются хорошим кормом для рыб на ранних этапах развития.
3.
Общая биомасса
зообентоса ильменя Горчичный составила 36,56 г/м2. Доминирующее
положение занимают представители семейства хирономид и гаммарид.
Следовательно, бентос
ильменя представлен видами, потребляемыми как молодью рыб, так и взрослыми
особями, что является предпосылкой вселения сюда рыб – бентофагов.
4. Ильмень Горчичный
представлен большим разнообразием планктонных и бентосных организмов,
необходимых для развития начальных звеньев и пищевых цепей, что является
большим плюсом возможности развития прудового хозяйства.
Список
литературы
1.
Абрамович Л.С.,
Аралина О.В., Василенко Т.П. Гидрохимические особенности солоноватоводных
прудов Присивашья // Материалы докладов 2 республиканской конференции
Украинского филиала ВГБО. – Киев: 1970. – с. 77-79.
2.
Абросов В.Н.
Проблема удобрения озер и связанные с ней вопросы // Известия ГосНИОРХ. – 1967.
–т. 64. – с. 61-68.
3.
Антипчук А.Ф.
Сезонная динамика численности микроорганизмов в солонаватоводных прудах юга
Украины // Рыбное хозяйство. – вып. 23. – Киев: 1972.-с. 43-46.
4.
Баранов И.В.
Гидрохимический режим и первичная продукция озер Морозовской группы Карельского
перешейка при сниженных нормах внесения минеральных удобрений // Известия
ГосНИОРХа, - 1972. – т. 79. – с. 29-44.
5.
Баранов И.В.
Временные методические рекомендации по минеральному удобрению малых озер. – Л.:
ГосНИОРХ, 1974.
6.
Баранов И.В.,
Салазкина А.Н. Химические и биологические методы повышения биопродуктивности
озер. – М: Пищевая промышленность, 1969. – 128 с.
7.
Бекин А.Г.
Влияние различных глубин на степень развития и интенсивность поедания бентоса
карпом в нагульных прудах // Материалы Всесоюзного совещания по кормовой базе.
– М.: 1973. – с. 53-55.
8.
Белевич Е.Ф.
«Ильмени Астраханского заповедника». Труды Астраханского Государственного
заповедника 1958 г. Выпуск 3.
9.
Белевич Е.Ф.
«Грунты подстепных ильменей дельты р. Волги» Труды Астраханского
Государственного заповедника 1958 г. Выпуск 3.
10.
Брюшков В.И.
«Западные подстепные ильмени». Труды Государственного океанографического
института. 1958.
11.
Галасун П.Т.,
Чижик А.К. Освоение высокоминерализованных водоемов и задачи науки //
Рыбоводство и рыболовство. – 1971. - № 3. – с. 14-15.
12.
Горбунов К.В.
Влияние зарегулирования Волги на биологические процессы в ее дельте и биосток.
– М.: 1976. – 218 с.
13.
Гордон Л.М.,
Эрман Л.А. Пути повышения эффективности товарного рыбоводства. – М.: Пищ.
пром., 1974. – 285 с.
14.
Горюнова В.Н.,
Никонова Р.С. Некоторые особенности выращивания двух – и трехлетков
растительноядных рыб и сазана в условиях дельты Волги / Труды ВНИИПРХа. – 1972.
– вып. 2. – с. 31-39.
15.
Еселевич В.Л.,
Еселевич М.И. В солоноватоводных водоемах на ирригационном стоке // Рыбоводство
и рыболовство. – 1970. - №3. – с. 16-17.
16.
Иванова М.Б.,
Умнов А.А. Способы определения продукции водных животных // Общие основы
изучения водных экосистем. – Л.: 1979. – с. 119-132.
17.
Китаев С.П.
Экологические основы биопродуктивности озер разных природных зон. М.: Наука,
1984. – 207 с.
18.
Комплексная
оценка экологической обстановки на Средней и Нижней Волге с использованием
природоохранного судна / Гуральник Д.Л. и др.. – Астрахань: 2000. – 276 с.
19.
Краснощек Г.П.
Фитопланктон рыбоводных прудов на юге Украины – естественная кормовая база для
толстолобиков // Рыбохозяйственное освоение прудов Молдавии. – Кишинев: 1974. –
с. 48-49.
20.
Кудерский Л.А.
Научные основы интенсификации рыбного хозяйства во внутренних водоемах страны
// рыбное хозяйство. – 1977. - №1. – с. 13-15.
21.
Кузьмин А.Н.,
Баллонов И.М. Фитопланктон // Методика изучения оиогеоценозов внутренних
водоемов. – М.: 1975. – с. 73-90.
22.
Лаврентьева И.В.
Первичная продукция прудов как показатель их рыбопродуктивности. Первичная
продукция морей и внутренних водоемов. – Минск: 1961. – с. 133-138.
23.
Малашкин Н.Н. О
методах направленного формирования ихтиофауны озер с целью получения товарных
уловов, близких к потенциальной продукции. – Труды Псковского отделения
ГосНИОРХа, 1976. – т. 2. – с. 19-24.
24.
Меретин А.В.
«Перспективы развития западно – подстепных ильменей Астраханской области» //
Рыбоводство и рыболовство// Москва 2002 Выпуск №1
25.
Материалы к
государственному докладу о состоянии окружающей природной среды РФ по
Астраханской области за 1999 год. – Астрахань: Из-во «ЦНТЭП», 2001. – 168 с.
26.
Новожилова М.И.,
Сокольский А.Ф., Горбунов К.В. Микрофлора и удобрения прудов аридной зоны СССР.
Алма-Ата: Наука, 1987. – 152 с.
27.
Оханов А.А.,
Сокольский А.Ф., Горбунов К.В. Из опыта организации озерных товарных рыбоводных
хозяйств в дельте Волги. – Астрахань: 1987.
28.
Петрова Т.Г.
Влияние разнокачественности рационов на рыбоводные и гематологические
показатели годовиков бестера в период зимнего содержания в садках на
отработанных водах ГРЭС // Методы индустриального рыбоводства. – М.: ВНИИПРХ,
1977. –с. 163-172.
29.
Пианка Э.К.
Эволюционная экология. – М.: Мир, 1981. – 400 с.
30.
Потенко Ю.С.
Продукция озерного бактериопланктона // Биологическая продуктивность эвтрофного
озера. – М.: Наука, 1970. – с. 81-88.
31.
Правдин И.Ф.
Руководство по изучению рыб. – М.: Пищ. Пром., 1966. – 374 с.
32.
Привольнее Т.И.
Отношение пресноводных проходных рыб к различной солености воды // Известия
ГосНИОРХа. – 1964. – т. 58. – с. 58-83.
33.
Природа и история
Астраханского края / Ушаков Н.М., Щукина В.П., Тимофеева Е.Г. и др.. – Астрахань:
Изд-во АГПУ, 1995. – 364 с.
34.
Решетников Ю.С.,
Попова О.А., Стерлигова О.П. Изменения структуры рыбного населения
эвтрофируемого водоема – М.: Наука, 1982. – 248 с.
35.
Руденко Г.П.
Характеристика малых озер и перспективы их рыбохозяйственного использования //
Известия ГосНИОРХа. – 1975. – т. 65. – с. 6-10.
36.
Руденко Г.П.,
Гвоздев М.А. Биопродукционаая характеристика и пищевые потребности рыб в озере
Кривое Псковской области // Известия ГосНИОРХа. – 1977. – т. 137. – с. 3-21.
37.
Рыжков Л.П. Биологические
и экономические основы развития озерных рыбных хозяйств // Материалы
Всесоюзного совещания по проектированию, строительству и эксплуатации озерных
рыбных хозяйств. – Петрозаводск: 1973. – с. 7-17.
38.
Рыжков Л.П.
теоритические аспекты развития озерного товарного рыбоводства // Тезисы
докладов XXI научной конференции по изучению и
освоению водоемов Прибалтики и Белоруссии. – т.1. – Псков: 1983. – с. 8-10.
39.
Сезонные явления
природы в западной части низовьев дельты Волги / Чуйков Ю.С. и др. // Природные
экосистемы дельты Волги Л.: 1984. – с. 121-128.
40.
Сокольская Н.И.
Формирование биопродуктивности водоемов дельты Волги и пути использование ее
закономерностей: Автореф. дис….канд. биол. наук. – М.: 1981. – 23 с.
41.
Сокольский А.Ф.,
Кокозова А.А., Мещеряков А.И., Горбунов К.В. Выращивание белуги в ильменях
дельты Волги // Формирование запасов осетровых в условиях комплексного
использования водных ресурсов. – Астрахань: 1986. –с. 328-329.
42.
Сокольский А.Ф.,
Мосолова Л.В., Николаева Т.А. Микробиологическая характеристика и фитопланктон
подстепных ильменей дельты Волги // Материалы Всес. Совещ. «Формирование и
регулирование естественной кормовой базы искусственных водоемов». – М.: 1973. –
с. 147-150.
43.
Сокольский А.Ф.,
Горбунов К.В., Меркулов А.Г. «Рыбохозяйственное использование ильменей дельты
Волги» / Тез. Докл. Всесоюз. конф. «Современное состояние и перспективы
развития прудового хозяйства» 1-3 декабря 1897 года. М.: 1987.
44.
Сокольский А.Ф.
«Современное состояние кормовой базы ильменей дельты р. Волга». / Биологические
ресурсы Каспийского моря. / Тез. Докл. первой международной конференции.
Сентябрь 1992 г. г. Астрахань.
45.
Сокольский А.Ф.,
Молодцов А.И., Степанова Т.Г., Курашова Е.К., Медная Л.И. «Состояние
биоресурсов и пути повышения продуктивности ильменей». / Биологические ресурсы
Каспийского моря и рационального их использования / по материалам 1993 г /
Астрахань 1994 г.
46.
Сокольский А.Ф.
«Биопродуктивность малых озер». Астрахань 1995.
47.
Сокольский А.Ф.
«Пути управления биоресурсами ильменей» / Ресурсы Каспийского моря: Тез. Докл. I Международной конференции. Октябрь
1992 г. Астрахань.
48.
Сокольский А.Ф.,
Еловенко В.Н. «Состояние запасов рыб в западных подстепных ильменях и
перспективы их промысла». // Первый конгресс ихтиологов России // Тез. Докл.
Астрахань 1997 г.
49.
Тютюник С.Н..
Конвейер производства рыбы – метод повышения рыбопродуктивности нагульных
прудов. – материалы конференции интенсификация рыбоводства на Украине. –
Херсон: 1974. – с. 79-80.
50.
Хлебович В.В.
Критическая соленость биологических процессов. – Л.: Наука, 1974. – 235 с.
51.
Цееб Я.Я. К
типологии солоноватых и соленых водоемов Крыма и характеристика их фауны //
Малые водоемы равнинных областей СССР и их использование. – М.: 1961. – с.
293-305.
52.
Чижик А.К.
рыбоводство в солоноватоводных прудах. – М.: Легк. и пищ. пром-сть, 1984. – 82
с.
|