Реферат: Аденилатциклазный сигнальный механизм
Реферат: Аденилатциклазный сигнальный механизм
АДЕНИЛАТЦИКЛАЗНЫЙ СИГНАЛЬНЫЙ МЕХАНИЗМ
ДЕЙСТВИЯ ПЕПТИДОВ ИНСУЛИНОВОГО СУПЕРСЕМЕЙСТВА У ПОЗВОНОЧНЫХ И БЕСПОЗВОНОЧНЫХ
Автореферат
диссертации на соискание ученой степени
доктора биологических наук
Санкт-Петербург 2007
Актуальность проблемы
Изучение гормональных сигнальных систем, участвующих
в регуляции жизненно важных для организма клеточных процессов, является одной
из актуальных проблем современной молекулярной эндокринологии и биохимии. К числу
систем, осуществляющих реализацию регуляторного действия веществ гормональной и
негормональной природы, относится аденилатциклазная сигнальная система (АЦС),
которая представлена в клетке сложным трансмембранным комплексом, состоящим, по
крайней мере, из трех молекулярных блоков. Необходимыми компонентами АЦС
являются: рецепторы, способные воспринимать внеклеточные сигналы,
гетеротримерные ГТФ-связывающие белки (G-белки) стимулирующего (Gs) или
ингибирующего (Gi) типа, состоящие из трех субъединиц – α, b, γ и обеспечивающие сопряжение между рецептором и третьим
компонентом системы - ферментом аденилатциклазой (АЦ), катализирующей образование
универсального внутриклеточного посредника – циклического аденозинмонофосфата
(цАМФ). При его участии осуществляется реализация целого ряда регуляторных
эффектов в клетке (пролиферация, дифференцировка, апоптоз, синтез белка и др.).
К настоящему времени в литературе накоплен
значительный объем данных, свидетельствующих об участии АЦС и цАМФ в
трансдукции сигналов группы гормонов не пептидной природы (серотонин,
адреналин, норадреналин и др.), осуществляющих свой эффект на клетку через
рецепторы серпантинного типа, семь раз пронизывающие мембрану. В рамках настоящего
исследования мы предприняли попытку выяснить возможность участия АЦС в
реализации действия пептидов инсулинового суперсемейства, обладающих
рецепторами тирозинкиназного типа, один раз пронизывающими мембрану клетки. До исследований,
проведенных нами, участие системы АЦС-цАМФ в реализации действия гормонов
инсулиновой природы практически отрицалось. В литературе имелись лишь отдельные
сведения о влиянии инсулина, бомбиксина, релаксина на активность АЦ (Pertseva
et al., 2003; Patel, 2004). Несмотря на успехи, достигнутые за последние
десятилетия в изучении молекулярных механизмов действия инсулина и
инсулиноподобных пептидов, многие аспекты плейотропного действия этого гормона
и родственных ему пептидов до сих пор остаются невыясненными (Pertseva et al.,
2003; Телкова, 2005). В связи с этим изучение ранее неизвестных молекулярных
механизмов действия пептидов инсулинового суперсемейства, насчитывающего в
настоящее время около 50 представителей, относится к числу актуальных проблем
современной эндокринологии. Согласно современным представлениям, инсулин и
родственные ему пептиды играют ключевую роль в регуляции ряда клеточных
процессов – клеточный рост, апоптоз, метаболизм. Эти пептиды имеют общее эволюционное
происхождение, так как возникли в ходе эволюции из общего анцестрального гена в
результате дупликации и последующей дивергенции образовавшихся генетических
линий, сохранив при этом структурное и функциональное сходство (Murray-Rust et
al., 1992; Chan et al., 1992).
К изучению участия АЦС в реализации
действия гормонов и ростовых факторов инсулиновой природы лаборатория
приступила в начале 90-х годов. Отправной точкой послужили данные, впервые полученные
нами, о способности инсулина и родственных пептидов активировать ГТФ-зависимым
образом АЦ в мышечных тканях млекопитающих и моллюсков (Plesneva et al., 1994).
Мы использовали эволюционный подход, предложенный Л.А. Орбели (1958) применительно
к эволюционной биохимии, который включал исследования в филогенезе, онтогенезе
и при патологии. Было изучено: 1) влияние на АЦС инсулина, инсулиноподобного
фактора роста 1 (ИФР-1) позвоночных и инсулиноподобного пептида (ИПП)
беспозвоночных (моллюск Anodonta cygnea; 2) влияние на АЦС пептидов в
тканях-мишенях животных разного филогенетического уровня (позвоночные – крысы,
птицы и беспозвоночные – моллюски); 3) действие пептидов инсулинового
суперсемейства на АЦС в онтогенезе (в тканях куриных эмбрионов разного возраста
и у цыплят); 4) действие пептидов инсулинового суперсемейства на АЦС при
экспериментальном диабете у позвоночных и беспозвоночных.
Цель работы. Доказать участие аденилатциклазной
сигнальной системы в реализации регуляторных эффектов инсулина, ИФР-1, ИПП
моллюска в клетке и расшифровать структурно-функциональную организацию АЦ
сигнального механизма их действия в тканях позвоночных и беспозвоночных, а
также установить роль АЦ сигнального механизма в регуляции фундаментальных
клеточных процессов – клеточный рост, апоптоз.
Задачи исследования
1. Исследовать действие инсулина, ИФР-1 и
ИПП, выделенного из висцеральных органов моллюска Anodonta cygnea (Русаков
и др., 1991), на АЦС в мышечных тканях позвоночных и беспозвоночных. Охарактеризовать
зависимость эффекта от времени и концентрации исследуемых пептидов, а также от
присутствия гуаниновых нуклеотидов для подтверждения вовлеченности в АЦ
сигнальный механизм гетеротримерных G-белков.
2. Исследовать структурно-функциональную
организацию АЦ сигнального механизма, опосредующего эффекты пептидов инсулинового
суперсемейства у позвоночных и беспозвоночных и выяснить последовательность
этапов передачи регуляторных сигналов этих пептидов на АЦС. С этой
целью: а) установить тип рецепторов и G-белков, вовлеченных в АЦ сигнальный
механизм действия пептидов, используя ингибиторы рецепторных тирозинкиназ и
метод АДФ-рибозилирования бактериальными токсинами; б) выявить участие фосфатидилинозитол-3
киназы (ФИ-3-К), используя специфичный ингибитор вортманнин; в)
идентифицировать изоформу протеинкиназы «С» (ПКС); используя ингибиторы ПКС и моноклональные
антитела к изоформам ПКС.
3. Исследовать участие АЦ сигнального
механизма действия инсулина и ИФР-1 в регуляции процессов клеточного роста и
апоптоза, исходя из гипотезы о важной роли цАМФ в регуляции фундаментальных процессов
в клетке (Перцева, 2000).
4. Исследовать функциональные нарушения в АЦ
сигнальном механизме действия пептидов инсулинового суперсемейства при
эндокринной патологии – сахарный диабет 1-го и 2-го типов.
Научная новизна
Впервые обнаружено стимулирующее действие
инсулина, ИФР-1 и ИПП моллюска A.cygnea на активность АЦ. Показано участие рецепторной тирозинкиназы
и установлена вовлеченность G-белков (Gi) и (Gs) типа в реализацию активирующего действия
этих пептидов на АЦ. Впервые показано, что в проявлении АЦ стимулирующих
эффектов инсулина и ИФР-1 участвуют ФИ-3-К и изоформы ПКС - ПКСz и возможно ПКСε.
В мышечных тканях позвоночных и
беспозвоночных животных обнаружен ранее неизвестный АЦ сигнальный механизм
действия инсулина и ИФР-1 и установлена его структурно-функциональная
организация, представленная в клетке следующей сигнальной цепью: рецептор тирозинкиназного
типа Þ Gi-белок (bγ-димер)
Þ ФИ-3-К Þ ПКСz (позвоночные) или ПКСε (беспозвоночные) Þ Gs-белок Þ АЦ Þ цАМФ Þ протеинкиназа «А» (ПКА) Þ эффекторные
системы. Этот механизм отличается по числу сигнальных блоков от известного АЦ
сигнального механизма действия гормонов, обладающих рецепторами серпантинного
типа, представленного в клетке следующей цепью: рецептор серпантинного типа Þ G-белок (Gi или Gs) Þ АЦ Þ цАМФ Þ ПКА Þ эффекторные
системы.
Следует отметить, что действие пептидов инсулиновой
природы на АЦ в мышечной ткани моллюска осуществляется через АЦ сигнальный
механизм, сходный с таковым позвоночных, но имеющий на пострецепторных этапах
трансдукции гормонального сигнала отличие на уровне ПКС. Исходя из результатов
нашего исследования, в АЦ сигнальном механизме действия пептидов инсулинового суперсемейства
у позвоночных принимает участие ПКСζ, а у беспозвоночных (моллюски), как
предполагается, – ПКСε.
Экспериментально подтверждена, выдвинутая
нами гипотеза, о важной роли АЦ-цАМФ в реализации регуляторного действия
инсулина и ИФР-1 на фундаментальные процессы в клетке. Показано участие АЦ-цАМФ
системы в способности ИФР-1 и инсулина стимулировать клеточный рост и ингибировать
апоптоз в культурах фибробластоподобных клеток.
Обнаружены нарушения в АЦ сигнальном
механизме действия инсулина при патологии (сахарный диабет 1-го и 2-го типов).
Теоретическое и практическое значение
работы
Теоретическое и практическое значение работы
определяется важной ролью инсулина и других пептидов инсулинового
суперсемейства в организме высших и низших животных. Обнаружение новых сигнальных
механизмов действия пептидов этой группы, в частности инсулина и ИФР-1,
расширяет современные представления о спектре сигнальных систем, участвующих в
регуляторном действии пептидов инсулинового суперсемейства.
Применение эволюционного подхода (изучение
ряда эволюционно-родственных пептидов и использование представителей
позвоночных и беспозвоночных) позволило выявить консервативность обнаруженного
АЦ сигнального механизма действия пептидов инсулиновой природы.
Данные, полученные на беспозвоночных, могут
быть полезны для понимания сигнальных механизмов действия пептидов инсулинового
суперсемейства у позвоночных и для разработки моделей эндокринной патологии у
человека (сахарный диабет) в рамках нового направления - «эволюционная биомедицина»
(Перцева, 2006).
Полученные данные о молекулярных
механизмах действия инсулина и ИФР-1 имеют фундаментальное значение и могут применяться
при чтении курса лекций в университетах и медицинских ВУЗах как в России, так и
за рубежом.
Результаты исследования имеют важное
практическое значение в плане выявления молекулярных основ этиологии и
патогенеза сахарного диабета, а также в создании новых подходов для диагностики
этого заболевания. Обнаруженный нами АЦ сигнальный механизм действия инсулина,
может служить основой для разработки биохимического теста, позволяющего
проводить диагностику нарушения отдельных звеньев в молекулярном механизме
действия инсулина.
Положения, выносимые на защиту
1. Впервые установлено, что пептиды
инсулинового суперсемейства (инсулин, ИФР-1 и ИПП моллюска Anodonta cygnea)
ГТФ-зависимым образом активируют АЦ в тканях позвоночных (млекопитающие, птицы)
и беспозвоночных (моллюски) животных.
2. Реализация АЦ активирующего действия пептидов
инсулиновой природы осуществляется через обнаруженный нами АЦ сигнальный
механизм, включающий следующую сигнальную цепь: рецептор-тирозинкиназа Þ Gi-белок (βγ-димер) Þ ФИ-3-К
Þ ПКСz Þ Gs-белок Þ АЦ.
3. С участием АЦ сигнального механизма,
генерирующего цАМФ, осуществляется регуляторное действие пептидов инсулиновой
природы на фундаментальные клеточные процессы - стимулируется клеточный рост и ингибируется
апоптоз.
4. При эндокринной патологии (сахарном
диабете 1-го и 2-го типов) нарушается функционирование АЦ сигнального механизма
действия гормонов инсулиновой природы в основном на уровне Gs-белка и его
сопряжения с АЦ.
5. Сходство структурно-функциональной
организации АЦ сигнального механизма действия пептидов инсулинового
суперсемейства у позвоночных и беспозвоночных животных свидетельствует об его
эволюционной консервативности.
Апробация работы
Основные результаты и положения работы
были представлены и доложены на следующих конференциях и съездах: 17-я, 19-я,
20-я, 21-я Конференции Европейского Общества Эндокринологов (Кордова, Испания,
1994; Нидерланды, 1998; Фаро, Португалия, 2000; Бонн, Германия, 2002); 4-й
Симпозиум по нейробиологии моллюсков (Амстердам, Нидерланды, 1994); Симпозиум
по инсулину, ИФР-1 и инсулиноподобным пептидам (Испания, Барселона,
1997); XXXIII Международный конгресс физиологов (Санкт-Петербург, 1997); 2-й
съезд Биохимического Общества РАН (Москва, 1997); XVII съезд физиологов России
(Ростов-на-Дону, 1998); конференция “Рецепция и внутриклеточная сигнализация” (Пущино,
1998); Съезд Биохимического общества Университета Глазго (Глазго,
Великобритания, 1999); 18-ый Международный конгресс биохимиков и молекулярных
биологов (Бирмингем, Англия, 2000); 3-я и 4-я Международные конференция по
релаксину и родственным пептидам (Брум, Австралия, 2000; Виоминг, США, 2004); XVIII
Съезд Физиологического Общества имени И.П. Павлова, Казань, 2001); XI, XII
и XIII Международные совещания по эволюционной физиологии (Санкт-Петербург, 1996;
2001; 2006); Международная Европейская конференция (Люксембург, 2002); Вторая конференция
“Эндокринная регуляция физиологических функций в норме и патологии”, посвященная
80-летию со дня рождения М.Г. Колпакова. (Новосибирск. Октябрь, 2002); 1-й
съезд Общества Клеточных Биологов (Санкт-Петербург, 2003); Физиологический
съезд (Екатеринбург, 2004); 1-й Съезд физиологов СНГ (Сочи, Дагомыс, 2005);
Публикации. По теме диссертации опубликовано 75 работ,
в том числе 36 статей в рецензируемых отечественных и международных изданиях.
Личный вклад автора – Экспериментальные данные
получены лично автором или при его непосредственном участии.
Структура и объем диссертации. Диссертация изложена на 259 страницах,
состоит из введения, обзора литературы, описания материалов и методов, изложения
результатов их обсуждения, заключения, выводов и списка литературы, включающего
285 источников. Работа иллюстрирована 43 рисунками и 35 таблицами.
Основное содержание работы
Объекты и методы исследования. Объектами исследования служили: 1)
представители позвоночных - крысы Ratus norvegicus линии Wistar; 2)
куры породы русская белая «Леггорн»; 3) представители беспозвоночных -
пресноводный двустворчатый моллюск Anodonta cygnea; 4) культура клеток
миобластов куриных эмбрионов; 5) фибробластоподобная культура клеток линии Swiss
3T3; 6) культура клеток, трансформированная из нормальных фибробластов линии
Е1А+сНа-ras (клетки, впадающие в апоптоз) и E1A+E1B (клетки, не впадающие в
апоптоз).
Методы. В работе использован широкий спектр физиологических,
биохимических и фармакологических методов:
- выделение фракций плазматических мембран
мышечной ткани позвоночных и беспозвоночных животных с использованием метода
дифференциального центрифугирования (Kidwai et. al., 1973 с нашими
модификациями);
- выделение частично очищенных мембранных
фракций культуры клеток миобластов куриных эмбрионов, фибробластоподобных
клеток линии Swiss 3T3, E1A+cHa-ras, E1A+E1B (Плеснёва и др., 1999; 2003);
- выделение фракции, содержащей примембранную
форму цАМФ-ФДЭ (Houslay, 1985).
- определение активности АЦ с
использованием радиоактивного субстрата АЦ реакции - [α32P]АТФ
(Salomon et al., 1974 с некоторыми модификациями);
- определение активности цАМФ-ФДЭ, с
использованием радиоактивного субстрата [3H]цАМФ (Ткачук и др.,
1978);
- АДФ-рибозилирование гетеротримерных G-белков
холерным и коклюшным токсинами (Pertseva et al., 1992);
- электрофорез в ПААГ
- иммуноблотинг с использованием
моноклональных антител для идентификации изоформы ПКСζ;
- определение ростстимулирующей активности
действия инсулина, ИФР-1, ЭФР и цАМФ по включению [14C] тимидина в
ДНК культуры клеток Swiss3T3 (Баркан
и др. 1992; Плеснёва и др., 1997; 1999);
- использование модели апоптоза на
трансформированных клетках, полученных из нормальных эмбриональных фибробластов
введением пары комплементирующих онкогенов E1A+cHa-ras, обладающих высокой
проапототической чувствительностью к удалению ростовых факторов (Bulavin et.al., 1999) и ее характеристика (Плеснёва и
др., 2003);
- оценка антиапоптотического действия
инсулина, ИФР-1 и цАМФ с использованием метода клоногенной выживаемости
культуры клеток E1A+cHa-ras (Плеснёва и др., 2003);
- определение активности ферментов
углеводного метаболизма – гликогенсинтетазы и глюкозо-6-фосфат-дегидрогеназы (Кузнецова,
1998; Кузнецова и др., 2004);
- определения содержания белков методом
Лоури;
- создания моделей экспериментального
диабета 1-го и 2-го типов у позвоночных (Ping et al., 1999 с нашими
модификациями) и диабетоподобного состояния у беспозвоночных (Плеснева и др.,
2006; Кузнецова и др., 2007) с использованием стрептозотоцина.
- статистические методы обработки данных
по программам (Statgraph и Anova).
Результаты исследования и их обсуждение
В работе представлены экспериментальные
доказательства активирующего действия инсулина, ИФР-1 и ИПП на АЦ, установлена
структурно-функциональная организация АЦ сигнального механизма в клетке и его
роль в регуляции пептидами инсулиновой природы клеточного роста и апоптоза.
Основные этапы работы состояли в
исследовании:
- действия пептидов инсулиновой природы на
активность АЦ при разном времени и разной концентрации in vitro и in vivo;
- участия примембранной цАМФ-ФДЭ в
функционировании АЦ сигнального механизма действия инсулиноподобных пептидов;
- звеньев АЦ сигнального механизма (от
рецептора до эффекторных систем);
- роли АЦ сигнального механизма, как в
регуляции клеточных процессов, так и его функционального применения в условиях
патологии;
Влияние in vitro инсулина, ИФР-1 и
ИПП на активность АЦ в тканях позвоночных и беспозвоночных и в культурах клеток
в разные временные сроки
Проведено исследование in vitro динамики
во времени АЦ активирующего эффекта инсулина, ИФР-1 и ИПП моллюска при
концентрации пептидов 10-8М и для сравнения эпидермального ростового
фактора (ЭФР), пептида не инсулиновой природы, обладающего рецептором
тирозинкиназного типа (Никольский и др., 1987). Показано, что в мембранной
фракции скелетных мышц крыс наибольшее выраженное активирующее действие исследованных
пептидов на АЦ проявляется через 2.5 мин. Активирующий АЦ эффект инсулина
составляет +236%. Эффект менее выражен при действии ИФР-1 (+201%), ЭФР (+186%)
и ИПП моллюска +124% (Рис. 1; Таблица 1).

Рис. 1.
Концентрация пептидов – 10-8М Активация АЦ пептидами (в%) по
сравнению с базальной активностью фермента, принятой за 100%, приведена в
тексте. Различия достоверны (р<0.05)
Таблица 1. Динамика во времени
действия ИФР-1 на активность АЦ в мембранной фракции скелетных мышц крыс
| Воздействия |
Активность АЦ (пкмоль цАМФ/мин/мг белка) |
| 2.5 мин |
5 мин |
10 мин |
| Без ИФР-1 |
71.2±4.8
(100%)
|
85.11±3.3
(100%)
|
101.20±9.24
(100%)
|
| + ИФР-1 |
214.31±12.1
(301%)
|
136.16±5.2
(160%)
|
96.14±4.6
(95%)
|
Примечание: В скобках - базальная активность
АЦ (без воздействий), принятая за 100% и активность АЦ (в%) при действии ИФР-1
В мембранной фракции культуры куриных
миобластов показано (рис. 2), что стимулирующий АЦ эффект инсулина через
2.5 мин составляет +256%, по сравнению с базальной активностью, принятой
за 100% и имеет сходство с данными, полученными на мембранной фракции скелетных
мышц крыс (+236%) (рис. 1).
В тканях моллюска A.cygnea, ИПП,
выделенный из висцеральных органов этого моллюска, оказывает наиболее
выраженное активирующее действие на АЦ (+422%), по сравнению с ЭФР (+221%),
ИФР-1 (+169%) и инсулином (+133%) (Рис. 3; Таблица 2). Следует отметить,
что активирующий АЦ эффект исследуемых пептидов в наибольшей степени
проявляется через 2,5 мин, через 5 мин снижается, а через 10 минут
практически отсутствует (Таблица 2; Рис. 3).
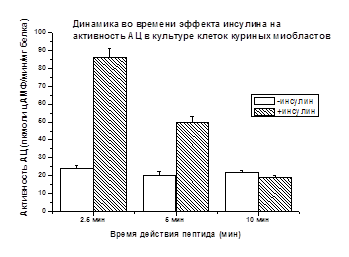
Рис. 2.
Концентрация инсулина 10-8М. Различия достоверны (р<0.05)
Таблица 2. Динамика во времени
действия ИФР-1 на активность АЦ в мембранной фракции гладких мышц моллюска
| Воздействия |
Активность АЦ (пкмоль цАМФ/мин/мг белка) |
| 2.5 мин |
5 мин |
10 мин |
| Без ИФР-1 |
110.21±10.8
(100%)
|
125.31±9.3
(100%)
|
106.04±6.6
(100%)
|
| + ИФР-1 |
259.92±11.4
(269%)
|
229.31±5.2
(183%)
|
103.81±5.8
(98%)
|
Примечание – как в таблице 1.
Таким образом, в мембранных фракциях,
выделенных из мышечных тканей исследуемых животных и культуры клеток куриных
миобластов, наблюдается закономерность проявления эффекта пептидов в зависимости
от времени действия. Инсулин, ИФР-1, ИПП и ЭФР оказывают активирующее действие
на АЦ в течение 2,5 и 5 мин с максимальным эффектом через 2.5 мин. АЦ
активирующий эффект исследуемых пептидов видоспецифичен. В скелетных мышцах
крыс ряд эффективности пептидов по их влиянию на АЦ имеет следующий порядок:
инсулин > ИФР-1 > ЭФР > ИПП, а в мышечных тканях моллюска - ИПП >
ЭФР > ИФР-1 > инсулин.

Рис. 3.
Концентрация пептидов – 10-8 М Активация АЦ пептидами (в %) по
сравнению с базальной активностью фермента, принятой за 100%, приведена в
тексте. Различия достоверны (р<0.05)
Влияние in vivo разных доз инсулина
на активность АЦ в тканях позвоночных и беспозвоночных в разные временные сроки
Обнаружив в опытах in vitro стимулирующее
влияние пептидов инсулинового суперсемейства на активность АЦ, мы исследовали влияние
инсулина, основного представителя инсулинового суперсемейства, на активность АЦ
в разных дозах и при разном времени действия в условиях in vivo. Эксперименты
проведены на фракциях мышечных мембран крыс, куриных эмбрионов и моллюсков
после введения инсулина (внутрибрюшинно или внутрижелточно) в дозах 0.05 нг/г-10
нг/г веса тела (вес крысы или куриного эмбриона, вес моллюска без раковины).
Доза, вводимого in vivo инсулина рассчитывалась с учетом концентрации
инсулина, используемой в опытах in vitro. Основанием для выбора дозы
служили данные литературы и результаты наших опытов in vitro. Было показано,
что в скелетных мышцах крыс стимулирующий АЦ эффект инсулина (+222%) выявляется
через 5 мин после введения гормона (1 нг/г), а через 15 мин он
снижается почти в три раза (67%) (Рис. 4). При более высокой дозе инсулина
(10 нг/г), стимулирующий АЦ эффект гормона через 5 мин выражен слабее (+124%),
а через 15 мин отсутствовал.
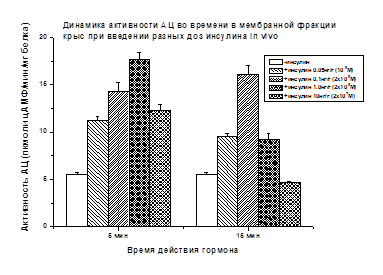
Рис. 4.
Активация АЦ пептидами (в %) по сравнению с базальной активностью АЦ, принятой
за 100%, приведена в тексте. Различия достоверны (р<0.05)
В экспериментах in vivo в
мембранных фракциях мышц куриных эмбрионов разного возраста обнаружен АЦ
активирующий эффект инсулина. У 10-дневных куриных эмбрионов он выявлялся уже через
5 мин, более четко выражен через 15 мин, а через 30 мин не проявлялся
(рис. 5).

Рис. 5. Различия достоверны (р<0.05)
У 17-дневных куриных эмбрионов выявлено дозозависимое
активирующее действие инсулина на активность АЦ через 5 минут (Рис. 6), которое
ослабевает через 15 и 30 мин после введения гормона.
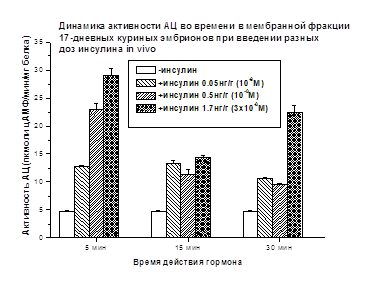
Рис. 6. Различия достоверны (р<0.05)
При введении моллюскам инсулина (0.5 нг/г
и 5.0 нг/г) активность АЦ возрастала через 5 мин после введения гормона на
+49% и +381%, соответственно, по сравнению с контролем, принятым за 100%. Через
20 мин влияние гормона (0.5 нг/г) на активность АЦ снижается (+22%) и
практически исчезает (+6%) через 40 мин (Рис. 7). При введении
инсулина (5 нг/г) моллюскам АЦ стимулирующий эффект гормона через 20 мин
составлял +110%, а через 40 мин +50% (рис. 7).
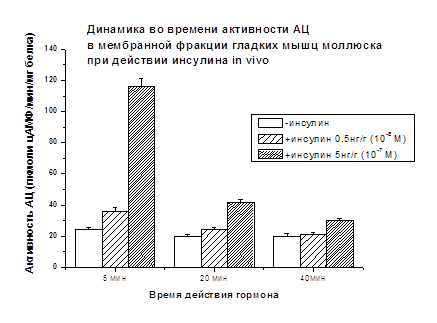
Рис. 7.
Светлые столбики - базальная активность АЦ. Заштрихованные столбики -
инсулин-стимулированная активность АЦ при разных дозах гормона. Различия
достоверны (р<0.05)
Из полученных нами данных следует, что отчетливое
влияние инсулина на активность АЦ в мышечных мембранах моллюска (Рис. 7) выявляется
при более высокой дозе инсулина, чем у млекопитающих и птиц. Это можно
объяснить тем, что у моллюска A.cygnea рецепторы к инсулину не обнаружены
(Лейбуш, Чистякова, 2003), а эффект инсулина может реализовываться через ИФР-1 -
подобные рецепторы или рецепторы ИПП моллюска, которые способны опосредовать активирующее
влияние инсулина на АЦ (Шпаков и др., 2005).
Следует отметить, что проявление АЦ
стимулирующего влияния инсулина и инсулиноподобных пептидов во времени имеет
сходство в опытах in vitro и in vivo. АЦ активирующий эффект
пептидов отчетливо выявляется при коротких сроках влияния пептида (2,5 и 5 мин)
при физиологических концентрациях (10-9-10-8М), с
увеличением времени действия эффект пептидов ослабевает или отсутствует.
АЦ активирующий эффект инсулина, ИФР-1 и
ИПП при разных концентрациях в условиях in vitro
Установив оптимальное время действия пептидов
инсулинового суперсемейства, при котором происходит активация АЦС (2.5 мин),
необходимо было выявить при каких концентрациях АЦ стимулирующий эффект
пептидов наиболее выражен. В разных тканях стимулирующий эффект пептидов инсулиновой
природы осуществляется через специфичные рецепторы, отличающиеся по сродству к пептиду
в гомологичных и негомологичных пептиду тканях.
Проведено исследование влияния инсулина,
ИФР-1, ИПП моллюска и ЭФР в течение 2.5 мин при разных концентрациях (10-12-10-6М)
в мембранных фракциях крыс, кур, моллюсков, а также во фракции, выделенной из
культуры клеток куриных миобластов.
В мембранной фракции скелетных мышц крыс
стимулирующий АЦ эффект инсулина, ИФР-1, ИПП обнаруживается при концентрациях -
10-11-10-7М (Рис. 8). Наиболее выраженный АЦ стимулирующий
эффект составляет: для инсулина +250% при 10-8М; для ИФР-1 +91% при
10-9М; для ИПП +111% при 10-8М; для ЭФР +190% при 10-10М.
(Рис. 8).
Можно отметить, что в диапазоне
концентраций 10-10-10-7М наиболее четко выражен
активирующий АЦ эффект инсулина, эффекты других исследованных пептидов
инсулиновой природы проявлялись слабее. АЦ стимулирующий эффект ЭФР проявлялся
при более низких концентрациях (10-11-10-10М).
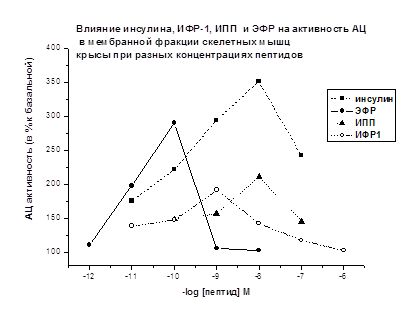
Рис. 8.
Базальная активность АЦ принята за 100%. Время действия пептидов 2.5 мин
Таблица 3. Влияние in vitro разных
концентраций инсулина на активность АЦ во фракциях мышечных мембран куриных
эмбрионов разного возраста и цыплят
| Объекты (возраст) |
10-сут
Эмбрионы
|
13-сут
Эмбрионы
|
16-сут
Эмбрионы
|
Цыплята
3-сут
|
| Воздействия |
Активность АЦ (пмоль цАМФ/мин/мг белка)
В скобках – Активность АЦ (в%) в присутствии инсулина,
по отношению к базальной активности, принятой за 100%.
|
|
Без
Инсулина
|
14.02±1.10
(100%)
|
3.90±0.45
(100%)
|
2.01±0.09
(100%)
|
8.52±0.41
(100%)
|
|
+инсулин
10-11 М
|
16.34±1.35
(117%)
|
4.21±0.34
(108%)
|
4.81±0.46
(239%)
|
10.11±0.55
(119%)
|
|
+ инсулин
10-10 М
|
39.14±1.93
(279%)
|
10.33±0.89
(265%)
|
5.28±0.30
(263%)
|
11.24±0.48
(132%)
|
|
+ инсулин
10-9 М
|
48.25±2.21
(344%)
|
15.92±1.24
(408%)
|
10.24±0.62
(509%)
|
14.54±0.82
(171%)
|
|
+ инсулин
10-8 М
|
19.87±1.44
(142%)
|
9.84±0.56
(252%)
|
9.92±0.47
(494%)
|
19.55±1.13
(238%)
|
|
+ инсулин
10-7 М
|
17.13±0.98
(122%)
|
4.93±0.45
(126%)
|
7.51±0.33
(374%)
|
22.21±1.37
(261%)
|
|
+ инсулин
10-6 М
|
17.90±1.12
(128%)
|
5.12±0.21
(131%)
|
1.95±0.22
(97%)
|
24.39±1.62
(286%)
|
Изучение АЦ стимулирующего эффекта
пептидов инсулинового суперсемейства и ЭФР в онтогенезе позволило выявить
следующие факты. Наиболее выраженный АЦ активирующий эффект инсулина (10-11М-10-6М
обнаруживается у куриных эмбрионов (10, 13, 16 суток) и цыплят (3 суток) при
концентрации гормона 10-9М, и составляет у 10-суточных эмбрионов +244%,
у 13-суточных +308%, у 16-суточных +409% (Таблица 3), а у 3-х суточных цыплят (+186%)
при более высокой концентрации 10-6М, что может быть связано с
переходом эмбрионов в постэмбриональный период, когда процессы роста и
дифференцировки заканчиваются.
Проведенные нами исследования действия
инсулина и ИФР-1 на активность АЦ в мембранной фракции, выделенной из культуры клеток
куриных миобластов (4-й день развития) показали, что наибольший АЦ стимулирующий
эффект инсулина (+352%) выявляется при концентрации 10-9М, а ИФР-1 (+289%)
при концентрации 10-10М (данные в диссертации).
В мембранной фракции мышц моллюска АЦ стимулирующий
эффект инсулина, ИФР-1, ИПП и ЭФР выявляется при концентрациях - 10-11-10-8М
(Рис. 9). Инсулин и ИФР-1 оказывают наиболее выраженное стимулирующее
действие на активность АЦ при концентрации 10-8М (+63% и +54%,
соответственно), а ИПП и ЭФР при 10-9М (+415% и +223%,
соответственно).
Таким образом, определен диапазон
концентраций, при которых проявляется АЦ активирующий эффект исследуемых
пептидов. Следует отметить видоспецифичность действия пептидов. Действие ИПП,
выделенного из висцеральных ганглиев моллюска A.cygnea, является более эффективным в ткани
моллюска и менее эффективным в тканях позвоночных.
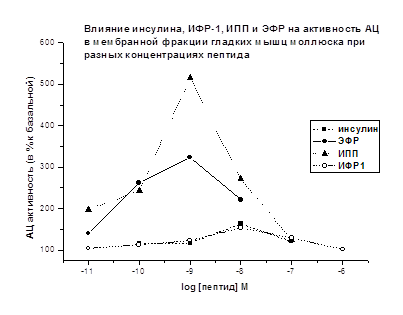
Рис. 9. Базальная активность АЦ принята за 100%. Время
действия пептидов – 2.5 мин.
Участие ц-АМФ-зависимой ФДЭ в механизме
действия пептидов инсулинового суперсемейства
Уровень цАМФ в клетке зависит не только от
АЦ – фермента, осуществляющего его синтез, но и от фосфодиэстеразы (ФДЭ) -
фермента, обеспечивающего его деградацию. Исследована динамика во времени активности
примембранной формы цАМФ-ФДЭ во фракции скелетных мышц кур. Эта изоформа ФДЭ
локализована на внутренней стороне мембраны и способна активироваться инсулином
(Houslay, 1985). Нами показано, что активность примембранной цАМФ-ФДЭ не
изменяется через 2.5 и 5.0 мин после действия инсулина, увеличивается на
+115% через 10 мин и на +179% через 20 мин, по сравнению с базальной
активностью фермента, принятой за 100% (Таблица 4).
Таблица 4. Влияние инсулина
(10-8М) на активность примембранной цАМФ-ФДЭ в скелетных мышц кур в
зависимости от времени
| Воздействия |
Активность цАМФ-ФДЭ (нмоль АМФ/мин/мг
белка) |
| 2.5 мин |
5.0 мин |
10.0 мин |
20.0 мин |
| Без инсулина |
3.37±0.5
(100%)
|
3.70±0.33
(100%)
|
3.56±0.61
(100%)
|
3.21±0.28
(100%)
|
| + инсулин |
3.06±0.21
(91%)
|
4.07±0.18
(110%)
|
7.65±0.23
(215%)
|
8.95±0.44
(279%)
|
Примечание: в скобках представлена
активность цАМФ-ФДЭ (в%) в присутствии инсулина и базальная активность фермента,
принятая за 100%
При исследовании действия разных
концентраций инсулина (10-9М-10-7М) обнаружено, что наиболее
выраженное действие гормона (+115%) на активность цАМФ-ФДЭ выявляется через 10 мин
при концентрации 10-8М (данные в диссертации).
Соотношение активности систем АЦ-цАМФ и
цАМФ-ФДЭ в реализации действия инсулина на клетку является важным моментом в
понимании внутриклеточных механизмов функционирования пептидов инсулиновой
природы.
При активации АЦ гормонами скорость
синтеза цАМФ начинает превышать скорость его деградации. Повышение уровня цАМФ
приводит к активации мишени его действия - ПКА. Сродство ПКА к цАМФ в 100-1000
раз больше, чем у ФДЭ. При усилении синтеза цАМФ происходит сначала насыщение
цАМФ-регуляторных центров ПКА, и лишь затем гидролиз цАМФ с участием ФДЭ.
Исходя из представленных нами данных,
активирующие эффекты инсулина на АЦ и цАМФ-ФДЭ четко разделены во времени. АЦ
активирующий эффект инсулина проявляется через 2.5 и 5 мин и отсутствует
через 10 мин. Между тем, цАМФ-ФДЭ активируется инсулином только через 10
минут инкубации с гормоном. Таким образом, активация цАМФ-ФДЭ наступает лишь
тогда, когда цАМФ как вторичный посредник выполнит свою функцию, достигнет
порогового уровня и осуществит свое активирующее действие на ПКА, а затем и на эффекторные
системы (Ткачук, 1983), что согласуется с нашими данными о действии инсулина на
активность АЦ и цАМФ-ФДЭ и данными литературы.
В связи с этим основное внимание будет
уделено исследованию АЦС, участвующей в осуществлении регуляторного действия
пептидов инсулинового суперсемейства на клеточные процессы.
Использование антиинсулиновой сыворотки для
доказательства специфичности АЦ стимулирующего эффекта инсулина
Для доказательства АЦ стимулирующего
действие инсулина, а не других пептидных гормонов, не относящихся к гормонам
инсулинового суперсемейства, была использована анти-инсулиновая сыворотка (АИС),
выработанная в лаборатории на инсулин млекопитающих, которая нейтрализует
действие инсулина и, как установлено нами подавляет стимулирующее действие
инсулина на АЦ. При добавлении нормальной сыворотки (без инсулина) к фракции
скелетных мышц крыс и гладких мышц моллюска, активирующий АЦ эффект инсулина
составляет +68% у крыс и +41% у моллюсков. При использовании АИС стимулирующий
АЦ эффект инсулина отсутствует у крыс (+7%) и моллюсков (-5%) (Табл. 5).
Полученные данные свидетельствуют о том,
что обнаруженное нами активирующее действие инсулина на АЦ в мышечных тканях
исследуемых объектов осуществляется именно инсулином и является специфичным при
действии этого гормона.
Таблица 5. Нейтрализация АЦ стимулирующего
эффекта инсулина в мембранных фракциях скелетных мышц крыс и гладких мышц
моллюска в присутствии антиинсулиновой сыворотки
| Воздействия |
Активность АЦ (пкмоль цАМФ/мин/мг белка) |
| Нормальная сыворотка |
Анти-инсулиновая сыворотка |
| Гладкие мышцы моллюска Anodonta cygnea |
| Без инсулина |
111.18±6.13 (100%) |
148.29±4.31 (100%) |
|
Инсулин 10-8М
|
156.76±8.54* (141%) |
158.67±8.16 (107%) |
| Скелетные мышцы крысы |
| Без инсулина |
97.34±4.58 (100%) |
115.82±6.49 (100%) |
|
Инсулин 10-8М
|
163.49±6.17* (168%) |
110.03±7.12 (95%) |
Примечание: приведены
данные из трех независимых экспериментов, повторенных три раза. Значения
представлены в виде среднего арифметического с учетом ошибки среднего. Различия
достоверны (р< 0.05 (*). В скобках – активность АЦ в %. Базальная активность
принята за 100%.
Следующая часть работы посвящена
расшифровке структурно-функциональной организации механизма АЦ активирующего
действия инсулина и ИФР-1.
В этой части работы мы поэтапно
исследовали звенья, которые могут быть вовлечены в реализацию действия
инсулиноподобных пептидов, начиная от рецептора и заканчивая эффекторными
системами, которые являются конечным звеном реализации сигнала.
Участие рецептора тирозинкиназного типа в
реализации АЦ стимулирующего эффекта пептидов инсулинового суперсемейства
Для доказательства участия рецептора
тирозинкиназного типа, характерного для пептидов инсулинового суперсемейства в
реализации АЦ стимулирующего эффекта пептидов инсулиновой природы, использовали
селективные ингибиторы рецепторных тирозинкиназ – тирфостин 47 и генистеин,
которые блокируют тирозинкиназную функцию рецептора инсулина и других пептидов
инсулиновой природы. Ингибиторы инкубировали с пробами в течение 15 мин
после чего в пробу добавляли пептиды (время действия 2.5 мин) при
концентрации, вызывающей максимальный АЦ стимулирующий эффект. Влияние этих ингибиторов
на активирующие АЦ эффекты инсулина, ИФР-1, ЭФР и для сравнения на эффект
изопротеренола (в случае позвоночных) и серотонина (в случае беспозвоночных),
которые также действуют активирующим образом на АЦ, но через рецептор
серпантинного типа, изучали in vitro во фракции мембранных препаратов
мышечной ткани крыс, культуры куриных миобластов, моллюсков.
У крыс в присутствии тирфостина 47
(1.25-5.0 мкМ) активирующий АЦ эффект пептидов снижался при действии инсулина
до 50%, при ИФР-1 до 65%, при ЭФР до 50% по отношению к максимальному эффекту
пептидов, принятому за 100% (Рис. 10). В присутствии генистеина (1.25-5.0
мкМ) снижение АЦ стимулирующих эффектов всех используемых пептидов у крыс было
более выражено. Эффект инсулина снижался до 40%, эффект ИФР-1 до 10%, эффект
ЭФР – до 18% (рис. 11). В тоже время у крыс стимулирующий эффект
изопротеренола на АЦ (10-6М) практически не изменялся в присутствии
тирфостина 47 и генистеина (0.5-20.0 мкМ).
В культуре клеток 4-х суточных куриных
миобластов тирфостин 47 (Рис. 12) и генистеин (данные представлены в
диссертации) ингибировали АЦ активирующий эффект инсулина до 43% и 38%,
соответственно. Однако, активирующее действие изопротеренола на АЦ в
присутствии ингибиторов тирозинкиназ (тирфостина 47 и генистеина) не изменялось
(р < 0.05). Это свидетельствует о том, что классические рецепторы
серпантинного типа не участвуют в механизме действия пептидов инсулиновой
природы.
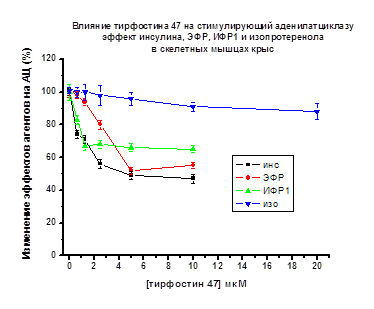
Рис. 10.
По оси ординат - снижение стимулирующего эффекта пептидов и изопротеренола на
АЦ (принятого за 100%) в присутствии тирфостина 47

Рис. 11.
По оси ординат - снижение стимулирующего эффекта пептидов и изопротеренола на
АЦ (принятого за 100%) в присутствии генистеина
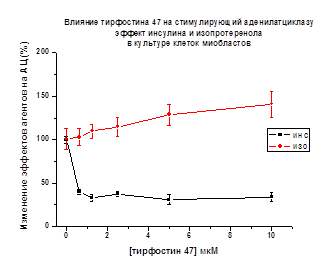
Рис. 12. По оси ординат - снижение стимулирующего
эффекта инсулина и изопротеренола на АЦ (принятого за 100%) в присутствии
тирфостина 47
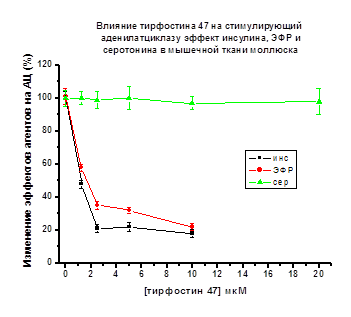
Рис. 13.
По оси ординат - снижение стимулирующего эффекта пептидов и серотонина на АЦ
(принятого за 100%)
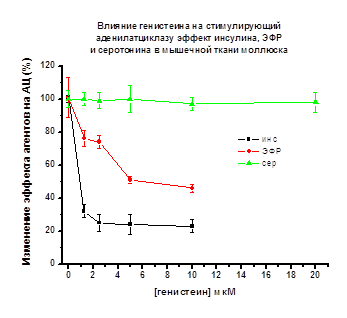
Рис. 14.
По оси абсцисс – концентрация генистеина в мкМ; по оси ординат - снижение АЦ
стимулирующего эффекта пептидов и серотонина (принятого за 100%) в присутствии
генистеина
У моллюсков наиболее выраженное снижение
АЦ стимулирующего эффекта пептидов наблюдалось в присутствии тирфостина 47 (2.5
мкМ). Эффект инсулина уменьшался до 20%, эффект ЭФР до 30-35%. (Рис. 13).
В присутствии генистеина АЦ стимулирующий эффект пептидов снижался до 25% в
случае инсулина (1.25 мкМ генистеин), и до 50-75% в случае ЭФР (5 мкМ генистеин)
(рис. 14).
Необходимо подчеркнуть, что у моллюсков A.cygnea активность АЦ увеличивается не в присутствии изопротеренола, а в
присутствии серотонина, реализующего свое действие также через рецепторы
серпантинного типа (Pertseva et al., 1992).
Исследование действия ингибиторов тирозинкиназ - тирфостина 47 и генистеина
(0.5-20.0 мкМ) на стимулирующий АЦ эффект серотонина в мембранной фракции
мышечных тканей моллюсков не выявило изменений в действии этого биогенного
амина на активность АЦ.
Таким образом, в мембранной фракции крыс,
моллюсков и культуры клеток куриных миобластов стимулирующее влияние
исследуемых пептидов на АЦ снижалось в присутствии тирфостина 47 и генистеина в
диапазоне концентраций - 1.25, 2.5, 5.0, 10, 20 мкМ во всех исследуемых объектах
(Рис. 10-14).
Стимулирующее действие изопротеренола
(крысы, куры) или серотонина (моллюски) на АЦ, реализуемое через рецепторы
серпантинного типа не изменялись в присутствии тирфостина 47 или генистеина в
исследуемых объектах (см. Рис. 10-14).
Можно заключить, что в мышечной ткани
млекопитающих (крысы), птиц (куры), культуре клеток куриных миобластов и в
мышцах моллюсков активирующее действие инсулина, ИФР-1, ЭФР на АЦ осуществляется
с участием рецепторов тирозинкиназного типа, специфичных для действия этих
пептидов. (Pertseva et al., 1996; Plesneva et al., 2003).
Участие
G-белков в реализации АЦ стимулирующего эффекта инсулина
Для доказательства участия G-белков в действии
инсулина на активность АЦ был применен широко распространенный подход, в котором
используется набор гуаниновых нуклеотидов, способных в разной степени либо стимулировать
ГТФ-азную активность G-белков в присутствии ГТФ и его аналогов - ГТФγS,
ГИДФ и тем самым активировать АЦ, либо ингибировать ГТФ-азную активность
G-белка в присутствии ГДФβS.
Было исследовано влияние ГТФ и ряда его
негидролизуемых аналогов на активность АЦ в присутствии и отсутствии гормона (Табл.
6).
Таблица 6. Влияние гуаниновых
нуклеотидов в отсутствии и присутствии инсулина на активность АЦ во фракции
мышечных мембран крысы и моллюска
| Воздействия |
Животные |
| Крыса |
Моллюск |
| Активность АЦ (%) |
| Контроль |
100±1.01% |
100±1.3% |
|
Инсулин (10-8М)
|
222±1.3%
(+122%)
|
186±1.8%
(+86%)
|
|
ГТФγS (10-5М)
|
242±1.4%
(+142%)
|
470±9.4%
(+370%)
|
|
ГИДФ (10-5М)
|
236±1.8%
(+136%)
|
269±4.9%
(+169%)
|
|
ГТФ (10-5М)
|
135±1.05%
(+35%)
|
163±5.1%
(+63%)
|
|
ГДФβS (10-5М)
|
95±1.2%
(-5%)
|
92±2.4%
(-8%)
|
| Инсулин + ГТФγS |
473±2.5%
(+373%)
[109%]
|
726±20.3%
(+626%)
[170%]
|
| Инсулин + ГИДФ |
399±8.2%
(+299%)
[41%]
|
441±12.3%
(+341%)
[86%]
|
| Инсулин + ГТФ |
277±10.1%
(+177%)
[20%]
|
279±8.4%
(+179%)
[30%]
|
| Инсулин + ГДФβS |
102±5.4%
(+2%)
[-17%]
|
105±4.3%
(+5%)
[-86%]
|
Примечание: в круглых скобках –
активирующий АЦ эффект используемых агентов в% по отношению к базальной
активности, принятой за 100%. В квадратных скобках – потенцирование эффекта
гормона в присутствии гуаниновых нуклеотидов в %.
Согласно представленным данным, ГТФγS,
ГИДФ, ГТФ стимулируют активность АЦ в мышечных мембранах крыс и моллюсков. При
совместном действии инсулина и гуаниновых нуклеотидов происходит усиление
(потенцирование) эффекта гормона по сравнению с аддитивным эффектом гормона и
гуаниновых нуклеотидов, действующих раздельно - в присутствии ГТФγS, ГИДФ
и ГТФ на +109%, +41% и +20% у крыс и на +170%, 86% и 30% у моллюсков (табл. 6).
ГДФβS же напротив снижает АЦ стимулирующий эффект инсулина как в мышцах
крыс, так и моллюсков.
Потенцирование
эффекта инсулина в присутствии ГТФγS, ГИДФ, ГТФ и отсутствие
потенцирующего эффекта в присутствии ГДФβS свидетельствует о вовлеченности
Gs-белков в АЦ сигнальный механизм действия пептидов инсулинового
суперсемейства.
Таблица 7. Влияние
коклюшного и холерного токсинов на базальную, инсулин- и ИФР1-стимулируемую
активность АЦ в скелетных мышцах крысы и моллюска A.cygnea
|
|
Активность АЦ (пкмоль цАМФ/мин/мг белка) |
| Воздействия |
Скелетные мышцы крысы |
Гладкие мышцы моллюска |
|
|
Без КТ |
+КТ |
Без КТ |
+КТ |
| Без пептидов |
39.7±3.4 |
48.5±2.0 |
63.2±4.1 |
69.1±9,6 |
|
|
(100%) |
(100%) |
(100%) |
(100%) |
| Инсулин |
67.9±3.6 |
48.2±2.7 |
200.5±14.4 |
74.2±7.6 |
|
10-9М
|
(171%) |
(99%) |
(317%) |
(108%) |
| ИФР-1 |
57.4±2.1 |
43.1±1.6 |
139.2±12.4 |
76.8±7.3 |
|
10-9М
|
(145%) |
(89%) |
(220%) |
(111%) |
|
|
Без ХТ |
+ХТ |
Без ХТ |
+ХТ |
| Без пептидов |
39.6±2.6 |
79.7±2.7 |
47.4±3.0 |
94.3±5.6 |
|
|
(100%) |
(100%) |
(100%) |
(100%) |
| Инсулин |
69.3±2.8 |
105.8±7.4 |
151.7±9.8 |
134.0±7.5 |
|
10-9М
|
(175%) |
(133%) |
(320%) |
(142%) |
| ИФР-1 |
56.7±4.2 |
106.2±6.5 |
100.0±5.4 |
122.6±8.8 |
|
10-9М
|
(143%) |
(133%) |
(210%) |
(130%) |
Примечание: В
скобках – активность АЦ в%. Активность АЦ без пептидов принята за 100%.
Для выяснения типов G белков, вовлеченных
в АЦ сигнальный механизм действия инсулина и ИФР-1 были использованы
бактериальные токсины (коклюшный и холерный), которые модифицируют α-субъединицы
Gi и Gs белков.
Коклюшный
токсин вызывает АДФ-рибозилирование αi-субъединицы Gi белка,
что ведет к потере его функциональной активности (Milligan, 1988; Reisine,
1990). Известно, что βγ-димер Gi белка обладает собственной
регуляторной способностью и может стимулировать активность ФИ-3-К. Обработка
мышечных мембран крысы и моллюска коклюшным токсином приводила к блокированию
АЦ стимулирующего эффекта, как инсулина, так и ИФР-1 (таблица 7), что можно
объяснить нарушением диссоциации гетеротримерного Gi белка на αi-субъединицу
и βγ димер в условиях действия коклюшного токсина.
Таким
образом, коклюшный токсин, предотвращая индуцируемую инсулином или ИФР-l
стимуляцию активности ФИ-3-К, реализуемую через βγ-зависимый
механизм, тормозит активацию АЦ.
Влияние холерного
токсина на мембраны приводит к блокаде ГТФ-азной активности αs-субъединицы
и тем самым переводит её в перманентно активированное состояние. В связи с этим
обработка мембран холерным токсином может повлечь за собой стимулирование
каталитической активности АЦ и наряду с этим ослабление регуляторных эффектов
гормонов, действие которых на АЦ осуществляется через Gs белок (Milligan, 1988;
Reisine, 1990). Обработка фракции мышечных мембран крысы и моллюска холерным
токсином приводит к 2х-кратному увеличению базальной активности АЦ и снижению
стимулирующего эффекта инсулина и ИФР-1 на активность фермента (таблица 7), что
полностью согласуются со сведениями литературы и указывает на вовлеченность Gs
белка в активацию АЦ с участием инсулина или ИФР-1.
Таким
образом, совокупность данных, полученных с использованием коклюшного и
холерного токсинов, указывает на участие как Gi, так и Gs белков в АЦ
сигнальном механизме действия инсулина и ИФР-l.
Участие
фосфатидилинозитол-3 киназы в реализации АЦ стимулирующего эффекта инсулина и
ИФР-1
Для выяснения
участия ФИ-3-К в АЦ сигнальном механизме действия пептидов инсулинового
суперсемейства (инсулина и ИФР-1) был использован специфический ингибитор этого
фермента - вортманнин. Инкубация мышечных мембран крысы и моллюска с
вортманнином (10-9–10-7М) несколько снижает базальную
активность АЦ (таблица 8). В отсутствии ингибитора инсулин и ИФР-1 отчетливо
стимулируют активность АЦ. Между тем, АЦ стимулирующий эффект инсулина и ИФР-1
снижается в зависимости от концентрации ингибитора (10-9–10-7М).
Ингибирующее действие вортманнина было наиболее выражено при концентрации 10-7М
(таблица 8). Установленные факты свидетельствуют об участии ФИ-3-К в АЦ
сигнальном механизме действия инсулина и ИФР-1 в мышечных тканях изучаемых объектов.
Таблица 8.
Влияние вортманнина (10–9М–10–7М) на стимуляцию ИФР-1 (10–8М)
и инсулином (10–8 М) активности АЦ в мембранной фракции скелетных
мышцах крыс и гладких мышц моллюска Anodonta cygnea
| Активность АЦ (пкмоль цАМФ/мин/мг белка) |
| объекты |
|
Крысы |
|
|
Моллюски |
|
| воздействия |
без пептида |
ИФР-l |
инсулин |
без пептида |
ИФР-l |
инсулин |
| без ворманнина |
21±1.6 |
38.2±1.0* |
41.4±2.3* |
17.8±1.0 |
41.1±2.6* |
24.5±1.0* |
|
+вортманнин
10–9М
|
17.9±2.0 |
9.4±1.3 |
9.7±1.4 |
15.8±2.0 |
14.6±1.3 |
14.2±0.4 |
|
+вортманнин
10–8М
|
16.5±2.3 |
8.6±1.3 |
8.4±1.3 |
14.6±2.3 |
13.9±0.8 |
11.6±1.0 |
|
+вортманнин
10–7М
|
13.2±1.9 |
6.3±0.9 |
6.8±0.8 |
14.3±0.9 |
13.8±1.8 |
7.4±0.5 |
Примечание:
значения активности АЦ в присутствии пептидов, достоверно отличающиеся от
активности фермента в отсутствии пептидов (р<0.05), отмечены
звездочкой.
Для проверки
гормоноспецифичности ингибирующего действия вортманнина на АЦ стимулирующие эффекты
инсулина и ИФР-1 были использованы изопротеренол и серотонин, гормоны неродственные
пептидам инсулиновой природы и реализующие действие через рецептор
серпантинного типа, не связанный с ФИ-3-К сигнальной системой. Результаты этих
опытов показали отсутствие влияния вортманнина на катехоламинчувствительную АЦ
сигнальную систему (данные приведены в диссертации). Этот факт указывает на
участие ФИ-3-К в, обнаруженным нами, АЦ сигнальном механизме действия инсулина
и ИФР-1.
Участие протеинкиназы С в АЦ сигнальном
механизме действия пептидов инсулинового суперсемейства
Для
доказательства участия ПКС в АЦ сигнальном механизме действия пептидов
инсулиновой природы использовали селективный блокатор ПКС – кальфостин (Yoing
et al., 2005). В отсутствии кальфостина АЦ активирующий эффект инсулина и ИФР-1
составлял +101% и +73% у крыс, и +78% и +86% у моллюсков соответственно, по
отношению к базальной АЦ, принятой за 100%. Как обнаружено нами, кальфостин (10-10–10-8М)
блокировал АЦ стимулирующие эффекты инсулина и ИФР-1 в мембранных фракциях мышц
крысы и моллюска. Наиболее выраженный ингибирующий эффект кальфостина
обнаруживается при концентрации 10-8М. В присутствии кальфостина (10-8М)
АЦ стимулирующий эффект инсулина и ИФР-1 снижался на 46% и 32% у крыс, и на 47%
и 50% у моллюсков, соответственно, по отношению к максимальному АЦ стимулирующему
эффекту пептидов, принятому за 100% (данные в диссертации).
Для
идентификации изоформы ПКС, участвующей в АЦ сигнальном механизме действия
пептидов инсулиновой природы использовали моноклональные антитела к ПКСζ,
которая по данным литературы является одним из участников реализации сигналов
инсулиновой природы.
Таблица 9.
Влияние антител к ПКСζ на АЦ активирующий эффект ИФР-1 (10-8М)
и инсулина (10-8М) в мышечных мембранах крысы и моллюска A.cygnea
|
|
Активность АЦ ( |
| Объекты |
|
Крысы |
|
|
Моллюски |
|
| Воздейтсвия |
Без пептида |
ИФР-1 |
Инсулин |
Без пептида |
ИФР-1 |
инсулин |
| без антител |
100±4.4 |
253±17 |
247±24 |
100±12 |
307±24 |
255±18 |
| АТ 1:1000 |
104±9.5 |
102±14 |
108±16 |
166±25 |
251±21 |
254±12 |
| АТ 1:100 |
98±8.2 |
95±12 |
103±5 |
163±18 |
327±27 |
237±20 |
| АТ 1:10 |
94±6.8 |
68±3.6 |
88±8 |
145±5 |
403±33 |
238±25 |
Активность АЦ
выражена в % к контрольным величинам, принятым за 100%.
Антитела к
ПКСζ почти полностью блокировали АЦ стимулирующий эффект инсулина и ИФР-1
в мышечных мембранах крыс (таблица 9). Таким образом, использование моноклональных
антител к ПКСζ показало, что эта изоформа фермента вовлечена в
стимулирующее действие инсулина и ИФР-1 на АЦ в скелетных мышцах крыс. Между
тем, в гладких мышцах моллюска эти антитела не оказывали блокирующего действия
на АЦ стимулирующий эффект инсулина и ИФР-1. Мышцы моллюска обладают иным
набором ПКС (Sossin et а1., 1996) и в АЦ стимулирующем действии этих пептидов
инсулинового суперсемейства, по-видимому, участвует другая изоформа ПКС,
близкая по своим свойствам к ПКСε из мозга позвоночных.
Таким образом, настоящее исследование
привело к обнаружению и расшифровке ранее неизвестного АЦ сигнального механизма
действия инсулина, ИФР-1 и ИПП моллюска в мышечных тканях позвоночных и
беспозвоночных животных. Этот механизм имеет принципиально сходную
структурно-функциональную организацию в случае инсулин- ИФР-1- и ИПП-компетентных
АЦ сигнальных систем. Он может быть представлен в клетке шестикомпонентным
сигнальным каскадом: рецептор-тирозинкиназа Þ
Gi-белок (βγ-димер) Þ
фосфатидилинозитол-3-киназа Þ протеинкиназа Сz Þ Gs-белок Þ
аденилатциклаза. АЦ является генератором внутриклеточного посредника – цАМФ, который
способен через активацию цАМФ-зависимой протеинкиназы “A” передавать
гормональный сигнал к различным эффекторным системам.
В плане изучения функциональной роли,
обнаруженного нами, АЦ сигнального механизма, генерирующего цАМФ, было
исследовано его участие в реализации регуляторного действия пептидов
инсулиновой природы на такие фундаментальные клеточные процессы, как клеточный
рост и апоптоз.
Участие АЦ
сигнального механизма в митогенном действии пептидов инсулиновой природы
Для изучения
митогенного действия пептидов инсулинового суперсемейства мы использовали
фибробластоподобную культуру клеток Swiss 3T3, любезно предоставленную нам из
банка культур Института цитологии РАН (Санкт-Петербург). Проведена функциональная
характеристика АЦ системы в культуре Swiss3T3 клеток. Установлено, что АЦ система в культуре этих клеток
Таблица 10.
Функциональные свойства АЦС культуры Swiss3T3 клеток
| Воздействия |
Активность АЦ
(пкмоль цАМФ/мин/мг белка)
|
Стимулирующий АЦ эффект в % |
| Базальная |
30.59±1.41 |
100% (базальная) |
|
NaF 10-2 М
|
178.91±4.15 |
585% (+485%) |
|
форсколин 10-5 М
|
99.26±3.21 |
324% (+224%) |
|
ГИДФ 10-6 М
|
111.20±3.48 |
364% (+264%) |
|
ГТФ 10-5 М
|
82.98±2.92 |
271% (+171%) |
|
ЭФР 10-9 М
|
168.31±4.75 |
550% (+450%) |
|
ИФР-1 10-9 М
|
102.14±3.34 |
334% (+224%) |
|
Инсулин 10-9 М
|
74.89±2.11 |
245% (+145%) |
В скобках –
активирующий АЦ эффект агентов гормональной и негормональной природы (в%), по
отношению к базальной активности АЦ, принятой за 100%, стимулируется
классическими негормональными активаторами АЦ - NaF, форсколином, ГИДФ, ГТФ, а
также инсулином, ИФР-1 и ЭФР (таблица 10).
Проведенные
нами исследования показали, что в культуре Swiss 3T3 присутствует АЦС с
функциональными свойствами, близкими к АЦС других клеток и тканей позвоночных,
которая способна к восприятию внеклеточных сигналов различной природы.
Оценив
функциональные свойства АЦ сигнальной системы культуры Swiss3T3 клеток, была
исследована способность инсулина, ИФР-1 и ЭФР индуцировать в культуре клеток
Swiss3T3 митогенный эффект, оцениваемый по включению [14C]-тимидина
в ДНК (Рис. 15-1). Показано, что инсулин (1000 нг/мл), ИФР 1 (50 нг/мл) и
ЭФР (10 нг/мл) стимулировали синтез ДНК (прирост от 100% до 250%).
Исследуемые
пептиды в тех же концентрациях оказывали стимулирующее влияние (2,5 мин)
на активность АЦ. (Рис. 15-2). Для подтверждения участия цАМФ в реализации
митогенного эффекта был использован его дибутириловый аналог цАМФ, обладающий
способностью проникать в клетку. Этот аналог, взятый при низких концентрациях
(10-12-10-9 М), вызывал четкий митогенный эффект, по
величине даже превосходящий аналогичный эффект ЭФР и ИФР-1. Эффект оценивали по
включению [14C]-тимидина в ДНК (Рис. 15-3).
Представленные
экспериментальные данные подтверждают нашу гипотезу (Перцева, 2000) об участии
АЦ сигнального механизма действия пептидов инсулинового суперсемейства и,
продуцируемого им цАМФ, в реализации митогенных процессов.
Участие
аденилатциклазного сигнального механизм в антиапоптотическом действии инсулина
и инсулиноподобного фактора роста 1
Нами подобрана модель апоптоза, включающая
клеточные линии, с разной степенью устойчивости к условиям, вызывающим
незапрограммированную гибель клеток (апоптоз). В экспериментах использовали
культуру клеток E1A+cHa-ras, обладающую высокой проапоптотической
чувствительностью к удалению ростовых факторов и действию ДНК-повреждающих
агентов («впадают в апоптоз») (Bulavin et al., 1999) и культуру клеток Е1А+Е1В
с высокой устойчивостью как к действию ДНК повреждающих агентов, так и к
удалению ростовых факторов из среды («не впадают в апоптоз»). В клеточных
культурах была охарактеризована чувствительность АЦС к действию инсулина и
ИФР-1 (Таблица 11).
Полученные результаты свидетельствуют о том, что
клетки культуры линий Е1А+сНа-ras и Е1А+Е1В способны отвечать на действие инсулина и ИФР-1
(10-8М) активацией АЦ, что
указывает на наличие в них рецепторов инсулина и ИФР-1, а также АЦС,
чувствительной к этим пептидам. Клетки сохраняют чувствительность АЦС к
инсулину и ИФР-1 как в среде с 10% сывороткой, так и в среде с 0.5% сывороткой.
Согласно данным литературы пептиды инсулинового
суперсемейства – инсулин и ИФР-1, а также цАМФ, образующийся в результате
активации АЦ, способны оказывать антиапоптотическое действие на клетки.
Показано, что инсулин (10-7М),
ИФР-1 (10-8М) и
дибутирил-цАМФ (10-9М)
оказывают ингибирующее влияние на апоптоз, вызванный удалением ростовых
факторов сыворотки, в культуре клеток E1A+cHa-ras (Плеснева, 2003). Оценка
антиапоптотического эффекта инсулина, ИФР-1 и дибутирил-цАМФ проводилась на
клетках культуры E1A+cHa-ras с использованием метода клоногенной выживаемости,
который относится к числу наиболее чувствительных способов тестирования
антиапоптотического действия агентов. Обнаружено, что культивирование
Таблица 11.
Влияние инсулина и ИФР-1 на активность АЦ в грубой мембранной фракции культур
клеток E1A+cHa-ras и E1A+E1B
| Условия |
Активность АЦ (пмоль цАМФ/мин/мг белка) |
| In vitro |
E1A+cHa-ras
(клетки, впадающие в апоптоз)
|
Е1А+Е1В
(клетки, не впадающие в апоптоз)
|
| Контроль |
Инсулин |
ИФР-1 |
Контроль |
Инсулин |
ИФР-1 |
| Среда + 10% сыворотка |
33.9±3.4
(100)
|
48.1±2.1
(142)
|
56.4±1.3
(166)
|
4.9±0.5
(100)
|
14.3±1.2
(292)
|
17.6±1
(359)
|
|
Среда + 0.5%
сыворотка (апоптоз)
|
95,9±3,4
(100)
|
137,0±9,0
(143)
|
181,6±8,2
(189)
|
26.7±0.5
(100)
|
61.4±1.2
(230)
|
86.3±2.1
(323)
|
| In vivo |
E1A+cHa-ras
(клетки, впадающие в апоптоз)
|
Е1А+Е1В
(клетки, не впадающие в апоптоз)
|
| Контроль |
Инсулин |
ИФР-1 |
Контроль |
Инсулин |
ИФР-1 |
|
Среда +10%
сыворотка
|
4.1±0.29
(100)
|
6.0±0.9
(146)
|
12.2±0.5
(297)
|
5.2±0.3
(100)
|
8.5±1.0
(163)
|
11.4±1.3
(219)
|
|
Среда + 0.5%
сыворотка (апоптоз)
|
11.0±1.2
(100)
|
17.2±1.2
(156)
|
24.8±2.2
(225)
|
5.8±0.6
(100)
|
10.0±0.7
(172)
|
13.6±0.6
(234)
|
|
|
|
|
|
|
|
|
Примечание. В
опытах in vitro гормоны (10-8М) добавляли прямо в пробу,
содержащую грубую мембранную фракцию клеток, для определения активности АЦ. Время
инкубации 2.5 мин. В опытах in vivo инсулин (10-7М) и
ИФР-1 (10-8М) добавляли к культурам клеток, время инкубации 5 мин.
Цифры в скобках - эффект гормонов в процентах к контролю, принятому за 100%.
Апоптоз индуцировали удалением ростовых факторов сыворотки из среды.
Трансформантов на среде без ростовых факторов
уменьшает число живых клеток, способных дать потомство в клональном посеве
минимум в 2 раза, по сравнению с клетками, культивируемыми в нормальных
условиях (среда+10% сыворотки).
Показано, что только 43% культуры клеток
«выживают» в условиях, когда клетки впадают в апоптоз (среда +0.5% сыворотки) по
сравнению с контролем (среда+10% сыворотка, принято за 100%). В экспериментах,
когда в среду с 0,5% сывороткой был добавлен инсулин, ИФР-1 или дибутирил-цАМФ
в указанных выше концентрациях, способность клеток давать жизнеспособное
потомство восстанавливалась до значений, составляющих около 70% (для инсулина:
77%, для ИФР-1: 67%, для дибутирил цАМФ: 66% по сравнению с контролем, принятым
за 100%).
Таким образом,
эксперименты показали способность инсулина и ИФР-1 через, обнаруженный нами, АЦ
сигнальный механизм и продуцируемый им цАМФ, оказывать митогенное и
антиапоптотическое действие в клеточных культурах.
Исследования
подтверждают участие АЦ сигнального механизма в реализации антиапоптотического
действия пептидов инсулинового ряда - инсулина и ИФР-1.
Функциональное
состояние АЦ сигнального механизма действия инсулина при сахарном диабете
На основе концепции молекулярных дефектов в
гормональных сигнальных системах как ключевых причин эндокринных заболеваний
(Перцева, 2004) исследовано функционирование АЦ сигнального механизма при
экспериментальном стрептозотоциновом сахарном диабете 1-го и 2-го типа у
позвоночных (крысы) и диабетоподобном состоянии у беспозвоночных (моллюски).
Диабет вызывали однократным введением (i.p.) стрептозотоцина (80 нг/г
веса животного). На 30-сут стрептозотоцинового диабета обнаружена гипергликемия
в крови крыс, в 4,2 раза превышающая уровень глюкозы у контрольных крыс.
Впервые выявлено увеличение уровня глюкозы в гемолимфе моллюсков (в 2,5 раза)
на 2-е и 4-е сутки развития диабетоподобного состояния. Обнаружено при диабете снижение
АЦ-стимулирующего эффекта инсулина и его потенцирования гуаниновыми
нуклеотидами у крыс и у моллюсков.
Инсулин независимый диабет (2-го типа) вызывали
введением новорожденным крысятам (1-2 сут) однократно стрептозотоцина (80 нг/г
веса животного). При этом типе диабета также возникают нарушения в АЦ
сигнальном механизме действия инсулина. АЦ стимулирующий эффект инсулина и
потенцирование не проявляется.
На основании полученных данных выявлены функциональные
дефекты в АЦ сигнальном механизме действия инсулина при диабете, в мышцах крыс
и моллюсков, затрагивающие дистальные звенья АЦ сигнального механизма на уровне
каталитического компонента – АЦ, Gs-белка и его сопряжения с АЦ.
Заключение
Настоящее исследование привело к
обнаружению и расшифровке ранее неизвестного АЦ сигнального механизма действия
инсулина, ИФР-1 и ИПП моллюска в мышечных тканях позвоночных и беспозвоночных
животных. Эти оригинальные данные расширяют современные представления о круге
сигнальных путей действия пептидов инсулинового суперсемейства и способствуют
формированию нового позитивного взгляда на вовлеченность АЦ сигнального механизма
в действие гормонов и ростовых факторов инсулиновой природы, осуществляемого
через рецепторы тирозинкиназного типа. До наших исследований в литературе
существовала точка зрения об участии АЦ сигнального механизма только в действии
гормонов, обладающих рецепторами серпантинного типа.
Важными представляются доказательства,
полученные в работе, о распространенности обнаруженного АЦ сигнального
механизма действия изученных пептидов в тканях как позвоночных, так и
беспозвоночных животных. Установлена принципиально сходная структурно-функциональная
организация инсулин-, ИФР-1- и ИПП-компетентной АЦ сигнальных систем. На
современном этапе наших исследований она представлена в клетке
шестикомпонентным сигнальным каскадом: рецептор-тирозинкиназа Þ Gi-белок (βγ-димер) Þ фосфатидилинозитол-3-киназа
Þ протеинкиназа Сz Þ Gs-белок Þ аденилатциклаза.
АЦ сигнальный механизм охватывает стадии от рецептора тирозинкиназного типа до
фермента – АЦ, являющейся генератором вторичного внутриклеточного посредника –
цАМФ. Образовавшийся цАМФ способен через активацию цАМФ-зависимой протеинкиназы
“A” передавать гормональный сигнал к различным эффекторным системам.
Открытый нами АЦ сигнальный механизм
действия пептидов инсулиновой природы по своей структурно-функциональной
организации наряду со сходством, обладает и существенными отличиями от
известных до сих пор гормональных АЦ сигнальных систем. Эти отличия сводятся:
- во-первых, к участию в одном АЦ
сигнальном механизме сразу дух типов гетеротримерных G-белков (Gs и Gi),
которые обычно вовлечены в разные сигнальные системы;
- во-вторых, к взаимодействию рецепторов
тирозинкиназного типа, специфичных для гормонов инсулиновой группы с Gi-белком,
выступающим в качестве донора βγ субъединиц, а не αi-субъединицы
как во многих других сигнальных системах.
- в-третьих, к большему числу,
составляющих его блоков (шесть компонентов, во всяком случае) по сравнению с
трехкомпонентным АЦ сигнальным механизмом действия биогенных аминов (рецептор серпантинного
типа Þ Gs или Gi-белок Þ АЦ);
За период, прошедший со времени
обнаружения нами АЦ сигнального механизма действия пептидов инсулинового
суперсемейства (Plesneva et.al., 1994; Kuznetsova et al., 1999; Plesneva et
al., 2001) в литературе появились данные, касающиеся отдельных его сигнальных
блоков, подтверждающие установленную нами структурно-функциональную организацию
этого АЦ сигнального механизма. Так показано, что рецепторы инсулина и ИФР-1
функционально сопряжены с Gi-белком (Kuemmerle, Murthy, 2001; Dupont et al.,
2003; Kreuzer et al., 2004). Также установлено участие ФИ-3-К и ПКСz и связь их с продукцией цАМФ в ряде сигнальных путей действия
пептидов инсулинового суперсемейства (Dessauer, Nguyen, 2005; Nguyen, Dessauer,
2005).
В плане изучения спектра функций,
обнаруженного нами АЦ сигнального механизма, генерирующего цАМФ, установлено
его участие в реализации регуляторного действия пептидов инсулиновой природы на
такие фундаментальные клеточные процессы, как клеточный рост (стимуляция) (Плеснева
и др. 1999) и апоптоз (ингибирование) (Плеснева и др., 2003), способствующие в
итоге выживанию клетки.
Таким образом, выдвинутая нами гипотеза (Перцева,
2001) о важной роли АЦ сигнального механизма в реализации регуляторного
действия инсулина и ИФР-1 на жизненно-важные клеточные процессы – клеточный
рост, апоптоз нашла подтверждение в наших исследованиях (Плеснева и др., 1999;
Плеснева и др., 2003).
Основываясь на эволюционном подходе Л.А. Орбели
ко всем изучаемым явлениям (Орбели, 1958) мы использовали комплекс методов эволюционной
физиологии применительно к изучению биохимических систем организма.
Исследование включало несколько аспектов: а) изучение трех эволюционно
родственных пептидов инсулинового суперсемейства – инсулина и ИФР-1
позвоночных, а также ИПП беспозвоночных (моллюска Anodonta cygnea); б) изучение
действия этих пептидов на АЦС в тканях-мишенях животных разного
филогенетического уровня (позвоночные – млекопитающие и птицы; а также беспозвоночные
– моллюск Anodonta cygnea); в) часть исследований (птицы) проведена в онтогенезе;
г) исследованы АЦ сигнальные механизмы действия пептидов инсулинового
суперсемейства и выявлены функциональные нарушения в них при патологии –
сахарном диабете.
В плане развиваемой в лаборатории
концепции (Перцева, Шпаков, 2004) молекулярных дефектов в гормональных
сигнальных системах как ключевых причин эндокринных заболеваний проведено
исследование функциональных нарушений в АЦ сигнальном механизме действия
инсулина и ИФР-1, возникающих при сахарном диабете. Впервые обнаружены дефекты в
этом сигнальном механизме на уровне каталитического компонента – АЦ, Gs-белка и
его сопряжения с АЦ, а также ослабление регуляторных метаболических эффектов
гормона у крыс с экспериментальным стрептозотоциновым диабетом 1-го и 2-го
типов.
Использование эволюционного подхода
позволило выявить не только существование АЦ сигнального механизма действия
ряда пептидов инсулинового суперсемейства у позвоночных и беспозвоночных, но и
обнаружить принципиальное сходство в его структурно-функциональной организации,
свидетельствующее об эволюционной консервативности этой сигнальной системы
Выводы
1. В мышечных тканях представителей позвоночных
(крысы) и беспозвоночных (моллюски) животных обнаружена чувствительная к влиянию
пептидов инсулиноподобного суперсемейства АЦС. Инсулин, ИФР-1 и ИПП моллюска
оказывают ГТФ-зависимое активирующее действие на АЦ. АДФ-рибозилирование бактериальными
токсинами позволило выявить участие G-белков как ингибирующего (Gi-белок), так
и стимулирующего (Gs-белок) типов в АЦ сигнальном механизме действия изученных
пептидов.
2. Установлено участие рецепторов тирозинкиназного
типа, в активирующем действии инсулина, ИФР-1 и ИПП моллюска на АЦС, ведущем к
образованию внутриклеточного посредника – цАМФ. Показано, что селективные
ингибиторы рецепторных тирозинкиназ – тирфостин 47 и генистеин, блокируют активирующее
действие исследуемых пептидов на АЦ в мышцах позвоночных и беспозвоночных.
3. Выявлено участие фосфатидилинозитол-3
киназы в АЦ сигнальном механизме действия пептидов инсулинового суперсемейства,
на что указывает блокирование активирующего влияния пептидов инсулиновой
природы на АЦ в присутствии специфического ингибитор ФИ-3-К – вортманнина.
4. Установлено участие протеинкиназы ПКСz идентифицированной с помощью моноклональных антител в реализации
АЦ стимулирующего действия пептидов инсулинового суперсемейства в мышечных
тканях позвоночных.
5. На основе совокупности полученных
данных, сделан вывод о существовании в клетках позвоночных, ранее неизвестного АЦ
сигнального механизма действия инсулина и ИФР 1, включающего сигнальную цепь:
рецептор-тирозинкиназа Þ Gi-белок(bγ) Þ фосфатидилинозитол-3-киназа Þ ПКСz Þ Gs-белок Þ
аденилатциклаза. Сходный механизм обнаружен в мышцах моллюска Anodonta cygnea,
отличающийся только изоформой ПКС.
6. Экспериментально подтверждена гипотеза
о важной роли АЦ сигнального механизма в реализации регуляторного действия инсулина
и ИФР-1 на фундаментальные клеточные процессы. Показана их способность
стимулировать клеточный рост (в культуре клеток Swiss3T3) и ингибировать апоптоз
(в культуре клеток Е1А+сНа-ras).
7. Обнаружение АЦ сигнального механизма
действия пептидов инсулинового суперсемейства природы в тканях-мишенях у
представителей как позвоночных, так и беспозвоночных животных, а также сходство
в его структурно-функциональной организации указывает на консервативность этого
сигнального механизма в эволюции.
8. При эндокринной патологии (сахарном
диабете 1-го и 2-го типов) у человека, а также у экспериментальных животных (позвоночных
и беспозвоночных) при стрептозотоциновой модели диабета обнаружены
функциональные нарушения в АЦ сигнальном механизме действия инсулина и ИФР-1,
локализованные в основном на уровне G-белка и его сопряжения с АЦ.
Публикации по теме диссертации
1. Pertseva M.N., Kuznetsova L.A.,
Plesneva S.A., Grishin A.V., Panchenko V.P. β-agonist-induced
inhibitory-guanine-nucleotide regulatory protein coupling to adenylate in
mollusk Anodonta cygnea foot muscle sarcolemma // Eur. J. Biochem.
Pharmacol. 1992. V. 210. P. 279-286.
2. Перцева М.Н., Плеснева С.А.,
Шпаков А.О., Русаков Ю.И., Кузнецова Л.А. Новые данные,
свидетельствующие об участии аденилатциклазной системы в механизме действия
инсулина и родственных пептидов // Доклады Академии наук. 1995. T. 342. №3.
C. 410-412.
3. Pertseva M.N.,
Plesneva S.A., Shpakov A.O., Rusakov Yu.I., Kuznetsova L.A.
Involvement of adenylyl cyclase signalling system in the action of insulin and
mollusc insulin-like peptide // Comp. Biochem. Physiol. 1995. V. 112. P. 689-695.
4. Перцева М.Н., Шпаков А.О.,
Плеснева С.А. Современные достижения в изучении сигнальных механизмов
действия инсулина и родственных ему пептидов // Журн. эвол. биохим. физиол.
1996.
T. 32. №3. C. 318-340.
5. Pertseva M.N.,
Plesneva S.A., Kuznetsova L.A., Shpakov A.O., Derkach K.V.
On the tyrosine kinase mechanism of the novel effect of insulin and
insulin-like growth factor-I: Stimulation of adenylyl cyclase system in muscle
tissues // Biochem. Pharmacol. 1996. V. 52. №12. P. 1867–1874.
6. Плеснева С.А., Баркан Р.С., Решетникова Г.Ф., Кузнецова Л.А., Перцева М.Н. Аденилатциклазный сигнальный механизм в
митогенном действии инсулина, инсулиноподобного и эпидермального факторов роста //
Доклады Академии наук, 1999. Т. 368. №6. С. 833-838.
7. Kuznetsova L.,
Plesneva S., Derjabina N., Omeljaniuk E., Pertseva M.N. On the
mechanism of relaxin action: the involvement of adenylyl cyclase signalling
system // Regulatory Peptides. 1999. V. 80. P. 33-39.
8. Плеснева С.А., Кузнецова Л.А., Омельянюк Е.В.,
Шпаков А.О., Перцева М.Н. Аденилатциклазный сигнальный механизм
действия релаксина // Журн. эвол. биохим. физиол. 2000. T. 36. №6. C.
562-568.
9. Кузнецова Л.А., Плеснева С.А.
Влияние биогенных аминов и полипептидных гормонов на активность
протеинкиназы А и аденилатциклазы в мышцах моллюска Anodonta cygnea //
Журн. эвол. биохим. физиол. 2001. Т. 37. №5.С. 395-400.
10. Plesneva S.A.,
Shpakov A.O., Kuznetsova L.A., Pertseva M.N. A dual
role of protein kinase "C" in insulin signal transduction via
adenylyl cyclase signaling system in muscle tissues of vertebrates and
invertebrates // Biochem. Pharmacol. 2001. V. 61. №10. P. 1277–1291.
11. Плеснева С.А., Шпаков А.О., Кузнецова Л.А.,
Перцева М.Н. Роль протеинкиназы С в регуляции процесса трансдукции
инсулинового сигнала через аденилатциклазный сигнальный механизм //
Российский Физиологический журнал им. И.М. Сеченова. 2001. T 87. №8. C.
1106–1117.
12. Шпаков А.О., Плеснева С.А.,
Кузнецова Л.А., Перцева М.Н. Исследование функциональной
организации нового – аденилатциклазного сигнального механизма действия инсулина //
Биохимия. 2002. T. 67. №3. C. 403-412.
13. Pertseva M.N.,
Shpakov A.O., Plesneva S.A., Kuznetsova L.A. A novel view
on the mechanisms of action of insulin and other insulin superfamily peptides:
involvement of adenylyl cyclase signaling system // Comp. Biochem. Physiol. 2003.
V. 134. №1. P. 11-36.
14. Шпаков А.О., Гурьянов И.А.,
Власова Е.Н., Корольков В.И., Кузнецова Л.А., Плеснева С.А.,
Власов Г.П., Перцева М.Н. Ингибирование синтетическими
катионными пептидами стимулирующего влияния гормонов на функциональную
активность аденилатциклазной сигнальной системы // Доклады Академии наук.
2003. T. 389. №1. C. 127-130.
15. Шпаков А.О., Плеснева С.А.,
Кузнецова Л.А., Леонтьева Е.А. Инсулинрегулируемая
аденилатциклазная сигнальная система в клеточной культуре фибробластов мыши
линии L // Журн. эвол. биохим. физиол. 2003. T. 39. №5. C. 438-444.
16. Плеснева С.А., Поспелова Т.В., Кузнецова Л.А.,
Быкова Т.В., Шпаков А.О., Перцева М.Н. Новая инсулинкомпетентная
аденилатциклазная сигнальная система как возможный механизм антиапоптотического
действия инсулина и инсулиноподобного фактора роста 1 // Доклады Академии
наук. 2003. T. 393. №4. C. 551-553.
17. Kuznetsova L.,
Shpakov A., Rusakov Yu., Plesneva S., Bondareva V., Pertseva M. Comparative
study of biological activity of insulin of lower vertebrates in the novel
adenylyl cyclase test-system // Regulatory Peptides. 2003. V. 116. №1-3. P.
81-86.
18. Кузнецова Л.А., Плеснева С.А.,
Шпаков А.О., Шарова Т.С. Стимулирующее влияние инсулина и
эпидермального фактора роста на активность протеинкиназы А,
глюкозо-6-фосфатдегидрогеназы и гликогенсинтетазы в скелетных мышцах кур в
эмбриогенезе // Журн. эвол. биохим. физиол.. 2004. T. 40. №4. C. 325-333.
19. Кузнецова Л.А., Плеснева С.А.,
Шпаков А.О., Бондарева В.М., Перцева М.Н. Инсулинрегулируемая
аденилатциклазная сигнальная система скелетных мышц крысы в условиях введения
инсулина in vivo и при инсулиновой недостаточности, вызванной
стрептозотоциновым диабетом // Журн. эвол. биохим. физиол. 2004. T. 40. №4.
C. 334-343.
20. Шпаков А.О., Шипилов В.Н.,
Кузнецова Л.А., Бондарева В.М., Плеснева С.А., Перцева М.Н. Реактивность
аденилатциклазной сигнальной системы нервных ганглиев моллюска Anodonta
cygnea к серотонину и адренергическим агонистам // Нейрохимия. 2004.
T. 21. №3. C. 190-197.
21. Шпаков А.О., Кузнецова Л.А.,
Плеснева С.А., Перцева М.Н. О вовлеченности фосфатидилинозитол-3-киназы
и протеинкиназы Сz в аденилатциклазный сигнальный механизм действия
релаксина в мышечных тканях крыс и моллюсков // Бюл. экспер. биол. мед.
2004. T. 138. №10. C. 420-423.
22. Шпаков А.О., Гурьянов И.А.,
Кузнецова Л.А., Плеснева С.А., Корольков В.И., Перцева М.Н.,
Власов Г.П. Использование С-концевых пептидов a-субъединиц G-белков для
исследования их функционального сопряжения с рецепторами биогенных аминов в
тканях крыс и моллюсков // Биол. мембраны. 2004. T. 21. №6. C. 441-450.
23. Шпаков А.О., Шипилов В.Н.,
Бондарева В.М., Кузнецова Л.А., Плеснева С.А., Русаков Ю.И.,
Перцева М.Н. Регуляторное действие родственных инсулину нейропептидов
моллюска Anodonta cygnea на функциональную активность аденилатциклазной
сигнальной системы // Нейрохимия. 2005. T. 22. №1. C. 28-37.
24. Шпаков А.О., Шипилов В.Н.,
Гурьянов И.А., Кузнецова Л.А., Бондарева В.М., Плеснева С.А.,
Перцева М.Н. Молекулярные механизмы регуляторного действия
биогенных аминов на функциональную активность аденилатциклазной сигнальной
системы в нервных ганглиях моллюска Anodonta cygnea // Доклады
Академии наук. 2005. T. 401. №6. C. 829-832.
25. Шпаков А.О., Кузнецова Л.А.,
Плеснева С.А., Гурьянов И.А., Перцева М.Н. Молекулярные
причины изменения чувствительности аденилатциклазной сигнальной системы
сердечной мышцы к биогенным аминам при экспериментальном стрептозотоциновом
диабете // Цитология. 2005. T. 47. №6. C. 540-548.
26. Шпаков А.О., Кузнецова Л.А.,
Плеснева С.А., Перцева М.Н. Молекулярные механизмы изменения
чувствительности аденилатциклазной сигнальной системы к биогенным аминам при
стрептозотоциновом диабете // Бюл. экспер. биол. мед. 2005. Т. 140. №9. С. 286-290.
|